
Di zarokatiyê de me hemûyan hîn kirin ku 5 hest hene: dîtin, çêj, deng, bîhn û dest. Çar hîsên destpêkê organên zelal, cihêreng, wek çav, tama, guh û poz bi kar tînin, lê tenê hesta laş tam çawa digihîje? Têkilî li ser tevahiya laş, hem li hundur û hem jî li derve tê ceribandin. Yek organek cûda tune ku berpirsiyariya hestiyariya destanê ye. Belê, li dora tevahiya laş receptorên piçûk, an dawiya nervê hene, ku li cîhê ku lê çêdibe têkilê hîs dikin û bi agahdariya li ser celebê desta ku çêbûye îşaretan dişîne mejî. Ji ber ku çêjekê li ser ziman tamê dişoxilîne, mekanoreceptor rijên di nav çerm û li ser organên din de ne ku hestên destdanê tespît dikin. Ew wekî têne zanîn mechanoreceptors ji ber ku ew ji bo tespîtkirina hestên mekanîkî an cûdahiyên zextê hatine çêkirin.
Contents
Rola Mechanoreceptors
Mirovek fam dike ku gava organê ku berpirsiyarê vedîtina wê hesta taybetî ye, peyamek dişîne mejî, ku organa bingehîn e ku hemî agahdariyan hildiberîne û birêkûpêk dike, ew hestek ceribandine. Ji hemî deverên laş bi têlên ku wekî neuron têne binav kirin ji mêjî re peyam têne şandin. Bi hezaran noyronên piçûk hene ku li hemî deverên laşê mirov şax dibin, û li ser dawiya gelek ji van noyronan mekanoreceptor hene. Ji bo ku nîşan bidin dema ku hûn destekê didin tiştekê çi diqewime, em ê mînakek bikar bînin.
Bifikirin ku mêşek li milê we dikeve. Zexta vê kêzikê, ew qas sivik, mekanoreceptorên li wê devera taybetî ya milê teşwîq dike. Ew mekanoreceptor li ser norona ku pê ve girêdayî ne peyamek dişînin. Neron bi tevahî rê bi mêjî ve girêdide, ku peyamê distîne ku tiştek bi laşê we li cîhê rastîn a mekanoreceptora taybetî ya ku peyam şandiye digire. Mejî wê bi vê şîretê tevbigere. Dibe ku ew ê ji çavan re bibêje ku li devera milê ku îmzeyê tespît kiriye binihêrin. Û dema ku çav ji mêjî re dibêjin ku li ser milê mêşek heye, mejî dibe ku ji dest re bibêje ku zû bifire. Bi vî rengî mekanoreceptor dixebitin. Armanca gotara li jêr ew e ku nîşan bide û hem jî bi hûrgulî li ser rêxistina fonksiyonel û diyarkerên molekulî yên mekanoreceptoran nîqaş bike.
Touch Sense: Organîzasyona Fonksiyonel û Determinantên Molekular ên Receptorên Mechanosensitive
Abstract
Mekanoreceptorên çerm di qatên cûrbecûr yên çerm de cih digirin, li wir ew cûrbecûr teşwîqên mekanîkî, di nav de firçeya sivik, dirêjbûn, lerzîn û zexta zirardar vedigirin. Ev cûrbecûr teşwîqan ji hêla cûrbecûr mekanoreceptorên pispor ve têne hev kirin ku bi rengek taybetî bersivê didin deformasyona çerm û van teşwîqan vediguhezînin strukturên mêjî yên bilind. Lêkolînên li ser mekanoreceptor û endikên hestiyar ên hestiyar ên genetîkî dest bi vedîtina mekanîzmayên hestiyariyê dikin. Xebata di vî warî de ji lêkolîneran re têgihiştinek bikêrhatî ya rêxistina dorpêçê ya ku di binê têgihîştina destikê de ye peyda kiriye. Kanalên îyonê yên nû wekî berendamên molekulên veguheztinê derketine û taybetmendiyên herikên bi mekanîkî yên dergehkirî têgihîştina me ya mekanîzmayên adaptasyona bi stimulasyonên taktîl çêtir kirine. Ev vekolîn pêşkeftina ku di karakterîzekirina taybetmendiyên fonksiyonel ên mekanoreceptoran de di çerm û kanalên îyonê yên porî û şirîn de ku têketinên mekanîkî tespît dikin û adaptasyona mekanoreceptorê çêdikin de ronî dike.
Keywords: mekanoreceptor, kanala mekanosensîv, êş, çerm, pergala somatosensor, dest
Pêşkêş
Têkilî tespîtkirina stimulasyona mekanîkî ye ku bandorê li çerm dike, di nav de teşwîqên mekanîkî yên bêguneh û zirardar. Ji bo zindîbûn û pêşketina memik û mirovan hestek bingehîn e. Têkiliya tiştên hişk û şilavan bi çerm re agahdariya pêwîst dide pergala nerva navendî ku destûrê dide lêgerîn û naskirina jîngehê û tevger an tevgera desta plankirî dide destpêkirin. Têkilî ji bo şagirtbûn, têkiliyên civakî û zayendîtiyê jî pir girîng e. Hestiya destgirtinê hesta herî kêm xeternak e, her çend di gelek şert û mercên patholojîk de dikare were guheztin (hîperesteziya, hîpoesteziya) jî.1-3
Bersivên destikê kodek pir rast a agahdariya mekanîkî vedigire. Mekanoreceptorên çermî di qatên cûrbecûr yên çerm de cih digirin, li wir ew cûrbecûr teşwîqên mekanîkî, di nav de firçeya sivik, dirêjbûn, lerzîn, guheztina por û zexta zirardar vedigirin. Ev cûrbecûr teşwîqan ji hêla cûrbecûr mekanoreceptorên pispor ve têne hevber kirin ku bi rengek taybetî bersivê didin deformasyona çerm û van stimulan vediguhezînin strukturên mêjî yên bilind. Neronên somatosensory yên çerm dikevin du koman: mekanoreceptorên tîrêjê nizm (LTMR) ku bertek nîşanî zexta bengê didin û mekanoreceptorên tîrêjê bilind (HTMR) ku bersivê didin teşwîqkirina mekanîkî ya zirardar. Laşên hucreya LTMR û HTMR di nav gangliyayên root ên dorsal (DRG) û gangliayên hestiyar ên kranial (gangliayên trigeminal) de dimînin. Fiberên nervê yên ku bi LTMR û HTMRs ve girêdayî ne li gorî leza tevgera wan a potansiyela çalakiya wan wekî fîberên A?-, A?- an C- têne dabeş kirin. Têlên C bêmyelîne ne û leza gihandina herî hêdî (~ 2 m/s) hene, lê A? û A? fiber bi sivikî û bi giranî myelinated in, bi rêzê ve leza gihandina navîn (~ 12 m / s) û bilez (~ 20 m / s) nîşan didin. LTMR di heman demê de li gorî rêjeyên wan ên adaptasyona ji stimulasyona mekanîkî ya domdar re wekî bersivên hêdî, an bilez ên adapteyî (SA- û RA-LTMRs) têne dabeş kirin. Ew ji hêla organên paşîn ên çermî yên ku ew dihejînin û teşwîqên wan ên bijarte ve têne cûda kirin.
Qabiliyeta mekanoreceptoran ji bo tesbîtkirina nîşanên mekanîkî bi hebûna kanalên îyonê yên mekanotransducer ve girêdayî ye ku bi lez hêzên mekanîkî vediguhezînin sînyalên elektrîkê û qada wergir depolarîze dikin. Ev depolarîzasyona herêmî, ku jê re potansiyela receptor tê gotin, dikare potansiyelên çalakiyê çêbike ku ber bi pergala nerva navendî ve belav dibe. Lêbelê, taybetmendiyên molekulên ku navbeynkariya mekanotransduction û adaptasyona hêzên mekanîkî dikin ne diyar in.
Di vê vekolînê de, em nihêrînek li ser taybetmendiyên mekanoreceptorê mammalan di têkila bêguneh û zerar de di çermê por û şêrîn de pêşkêş dikin. Di heman demê de em zanyariyên vê dawiyê yên di derbarê taybetmendiyên herikên mekanîkî-dergehkirî de jî dinirxînin ku hewl didin ku mekanîzmaya adaptasyona mekanoreceptorê rave bikin. Di dawiyê de, em pêşkeftina vê dawiyê ya ku di naskirina kanalên ion û proteînên têkildar de berpirsiyar in ji bo hilberîna herikên mekanîzmayî dinirxînin.
Têkiliya Bêbawer
LTMR-yên ku bi pelika porê ve girêdayî ne
Pûçikên por organên piçûk ên ku tîrêjên tîrêjê çêdikin temsîl dikin. Fiberên ku bi pelikên porê ve girêdayî ne bersivê didin tevgera por û rêwerziya wê bi avêtina trênên potansiyelên çalakiyê di destpêkê û rakirina stimulusê de. Ew bi lez receptoran adapte dikin.
Pisîk û kevroşk. Di kirasê pisîk û keroşkê de, pelikên por dikarin li sê celebên porê porê bêne dabeş kirin, porê Down, porê Guard û Tylotrichs. Porên berjêr (binê por, hirî, velûs)4 mûyên herî pir, kurttirîn û herî tenik in. Ew pêl, bêreng in û bi komên ji du-çar mûyan ji dirûvek hevpar a di çerm de derketine. Porên cerdevanan (monotrichs, overhears, tophair)4 hinekî kilkirî ne, yan pigmentî ne yan jî bêpîgment in û bi tenê ji devê pelikên wan derdikevin. Tîlotrîx mûyên herî hindik in, yên herî dirêj û stûr in.5,6 Ew pîgmented an bêpîgment in, carinan herdu jî û bi tena serê xwe ji folîkulek ku bi lepek damarên xwînê yên kapilar ve hatiye dorpêçkirin, derdikevin. Têlên hestiyar ên ku ji pelika porê re peyda dikin li binê girêza sebaceousê cih digirin û ji A? an fîberên A?-LTMR.7
Di nêzîkatiya mêşa porê xwarê de, tam li jêr asta gêrika sebaceous zengila dawiya pilo-Ruffini lanceolate heye. Van endên nervê yên hestî di qursek spiral de li dora porê porê di hundurê tevna girêdanê de ku pelika porê pêk tîne de cih digirin. Di hundurê pelika porê de, dawiya nervê ya belaş jî hene, hin ji wan mekanoreceptoran ava dikin. Pir caran, korpûsûlên destan (binihêrin çermê zelal) dora stûyê folîkula tylotrich dorpêç dikin.
Taybetmendiyên endikên nervê yên mîlînkirî yên di çermê porê pisîk û keroşkê de di heyama 1930-1970 de bi giranî hatine vekolîn (vedîtin li Hamann, 1995).8 Bi balkêşî, Brown û Iggo, 772 yekîneyên bi fiberên nervê yên aferent ên myelinated ên di demarên saphenous de lêkolîn kirin. û keroşk, di sê celeb receptoran de bersivên ku bi tevgerên mûyên Down (receptorên tîpa D), porê cerdevan (receptorên tîpa G) û porê Tylotrich (reseptorê tîpa T) ve girêdayî ne. di receptora Bi Bilez Adapkirinê ya tîpa I (RA I) de ji ber dijberiya receptorê Pacinian bi navê RA II. Mekanoreceptorên RA I leza stimulasyona mekanîkî tespît dikin û sînorên tûj hene. Ew guhertinên germî nabînin. Burgess et al. di heman demê de receptorek zeviyê ya ku bi lez adapte dibe jî destnîşan kir ku bi rengek çêtirîn bersivê dide lêdana çerm an tevgera çend poran, ku ji teşwîqkirina dawiya pilo-Ruffini ve hatî girêdan. Yek ji bersiva pelika porê ji çalakiya fîbera C re nehatiye veqetandin.9
Mice. Di çermê porê piştê yê mişkan de, sê cureyên sereke yên pelikên porê hatine diyarkirin: zigzag (nêzîkî 72%), awl/auchene (nêzîkî 23%) û guard an tylotrich (nêzîkî 5%).11-14 Zigzag û Awl/ Pûçikên porê auchenne şêlên porê ziravtir û kurttir çêdikin û bi yek gêrika sebaceous ve girêdayî ne. Porên cerdevan an tylotrich ji celebên porê porê herî dirêj in. Ew bi ampûlek porê mezin a ku bi du glandên sebaceous ve girêdayî ne têne destnîşan kirin. Porên cerdevan û awî/auchene bi şêweyek dubarekirî, bi rêkûpêk veqetandî têne rêz kirin lê ku porên zigzag bi zexmî deverên çerm ên li dora du celebên porê mezin ên porê vedihewînin [Fig. 1 (A1, A2 û A3)].
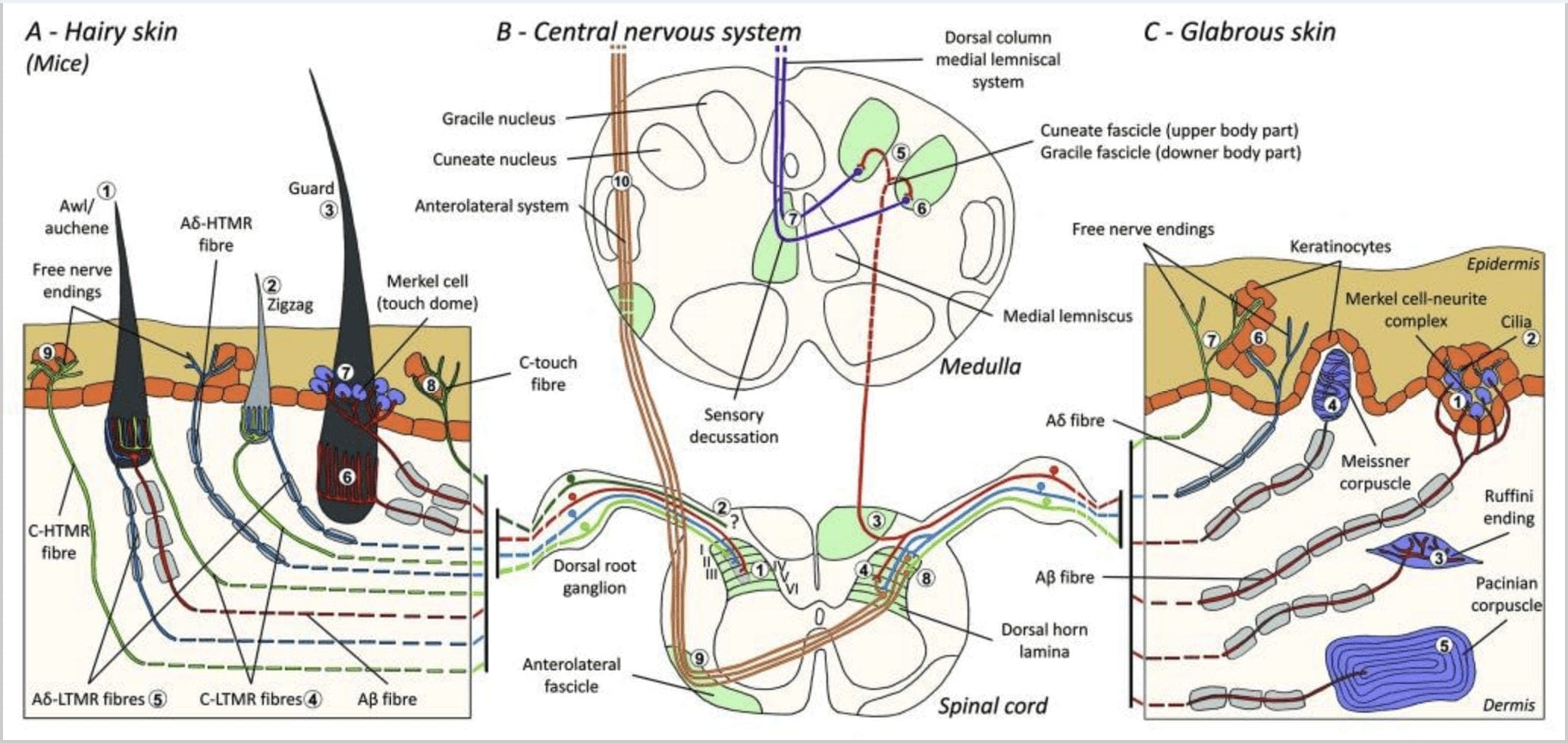
1. Rêxistin û pêşbîniyên mekanoreceptorên çerm. Di çermê porî de, firçeya sivik û destdan bi giranî ji hêla bêhnvedana li dora pelikên porê ve têne tespît kirin: awl/auchenne (A1), zigzag (A2) û cerdevan (A3). Porên awl/auchene sê caran ji hêla C-LTMR-ên lanceolate (A4), A?-LTMR û A? bi lez adapteyî-LTMR (A6). Pûçikên porê zigzag şêlên porê kurttir in û hem ji hêla C-LTMR (A4) û hem jî ji hêla A ve têne nerazî kirin? -LTMR dawiya lanceolate (A5). Pîskên porê cerdevan ên herî dirêj ji hêla A? Bi lez adapteyî-LTMR dawiya lanceolate dirêj (A6) û bi A re têkildar in? hêdî hêdî adaptekirina-LTMR-ya dawînên qubeya desta (A7). Pêşniyarên navendî yên van hemî fiberan bi pelikên cihêreng, lê bi qismî hevgirtî yên stûyê piştê yê mêjî (C-LTMR li lamina II, A?-LTMR li lamina III û A?-LTMR di lamina IV û V de) bi dawî dibin. Pêşniyazên LTMR-ê yên ku heman pelikên porê an jî cîranê xwe diherikînin têne rêz kirin da ku stûnek teng di stûna piştê ya stûnê de (B1 bi gewr) ava bikin. Tenê di çermê porî de, binekomek ji fîberên C-yê ku dawiya wê ya bêpere ye, epîdermisê nerazî dike û bersivê dide pêwendiya xweş (A8). Van fîberên C-touch bersivê nadin pêwendiya zirardar û rêwîtiya riya wan hîn jî nayê zanîn (B2). Di çermê zelal de, pêwendiya bêguneh ji hêla çar celeb LTMR ve tê navgîn kirin. Kompleksa hucre-neurite ya Merkel di tebeqeya bingehîn a epidermis (C1) de ye. Ev mekanoreceptor ji lihevhatina di navbera gelek hucreyên Merkel û termînalek nervê ya mezinbûyî ya ji yek A pêk tê? mûyik. Hucreyên Merkel pêvajoyên mîna tilikê ku bi keratinocytes (C2) re têkilî daynin nîşan didin. Dawiya Ruffini di dermisê de cih digire. Ew qertafek hestî ya bi şiklê cigarê tenik e ku bi A? fiber (C3). Corpuscle Meissner girêdayî A? dawiya nervê ye û di papillae dermal de cih digire. Ev mekanoreceptorê vegirtî ji şaneyên piştgirî yên pakkirî pêk tê ku wekî lameleyên horizontî yên ku ji hêla tevna girêdanê (C4) ve hatine dorpêç kirin pêk tê. Corpuscle Pacinian mechanoreceptor kûrtir e. Yek yek A? dawîya nervê ya bêmielîn di navenda vê korpuskula hêşînayî ya mezin a ku ji lameleyên hevsengî hatiye çêkirin bi dawî dibe. Pêşniyarên van fîberên A?-LTMR di mêjûya spî de li du şaxan têne dabeş kirin. Şaxa navendî ya sereke (B3) di mêjûya spinal de di dorsalê ya heman alî de hildiweşe û li ser astê medullayê fasîkûlên cuneate an gracil (B5) çêdike ku aferentên seretayî sinapsa xwe ya yekem çêdikin (B6). Neronên duyemîn decussasyonek hestî (B7) çêdikin da ku rêçek li ser lemnîskûsa navîn ava bikin ku di nav stûna mêjî de berbi mêjiyê navîn, bi taybetî di talamusê de, hilkişe. Şaxê duyemîn ê LTMR di qurmê dorsalê de di lamina II, IV, V de bi dawî dibe û bi veguheztina êşê (B4) re têkildar dibe. Têkiliya xedar bi dawiya nerva azad a di epîdermisê ya çermê pirç (A9) û çermê şil (C7) de tê tespît kirin. Van mekanoreceptor dawiya A?-HTMR û C-HTMR ne ku bi keratinocytes cîran (C6) re di têkiliyek nêzîk de ne. A?-hTMR di lamîna I û V de bi dawî dibe; C-HTMR di lamina I û II (B8) de bi dawî dibe. Di asta stûyê dorsalê de, aferentên seretayî HTMR bi noyronên duyemîn re sinapsan çêdikin ku ji xeta navîn derbas dibin û di fasîkula anterolateral (B9, B10) de berbi avahiya mejî ya bilindtir ve hildikişin.
Di van demên dawîn de, Ginty û hevkarên xwe ji bo dîtina organîzasyona endokên aksonal ên derdor û navendî yên LTMR-yên di mişkan de berhevokek nîşankirina molekul-genetîk û nêzîkatiyên şopandina paşverû ya somatotopîk bikar anîn. ji hêla sê celebên porê porê ve têne derxistin û bi çalakiyên yekbûyî yên yekbûyî yên A?-, A?- û C- tîrêjên dorsalê ve têne veguhestin.
Wan destnîşan kir ku nîşankirina genetîkî ya tîrosîn hîdroksîlase ya pozîtîf (TH+) neyronên DRG nifûsa noyronên hestî yên nepeptiderjîk, bi pîvana piçûk diyar dike û rê dide dîtina dawiya dorhêla C-LTMR di çerm de. Ecêb e, şaxên axoneal ên C-LTMR-yên takekesî hatin dîtin ku bi çîçikan ve çêdibin û dawiya lanceolate dirêjî ku ji nêz ve bi zigzag (80% ji dawiyan) û awl/auchene (20% ji dawiyan) ve girêdayî ne, lê ne pelikên porê tylotrich pêk tînin [Fig. 1 (A4)]. Demek dirêj hate fikirîn ku dawiya lanceolate ya dirêjî bi taybetî girêdayî A?-LTMR-yan e û ji ber vê yekê nedihat hêvîkirin ku dawiya C-LTMR-yên dirêjî yên lêdanê çêbike.15 Van C-LTMR-yên hanê li gorî adaptasyona hêdî û bilez xwedan adaptasyonek navîn in. mekanoreceptorên myelinated [Fig. 2 (C1)].
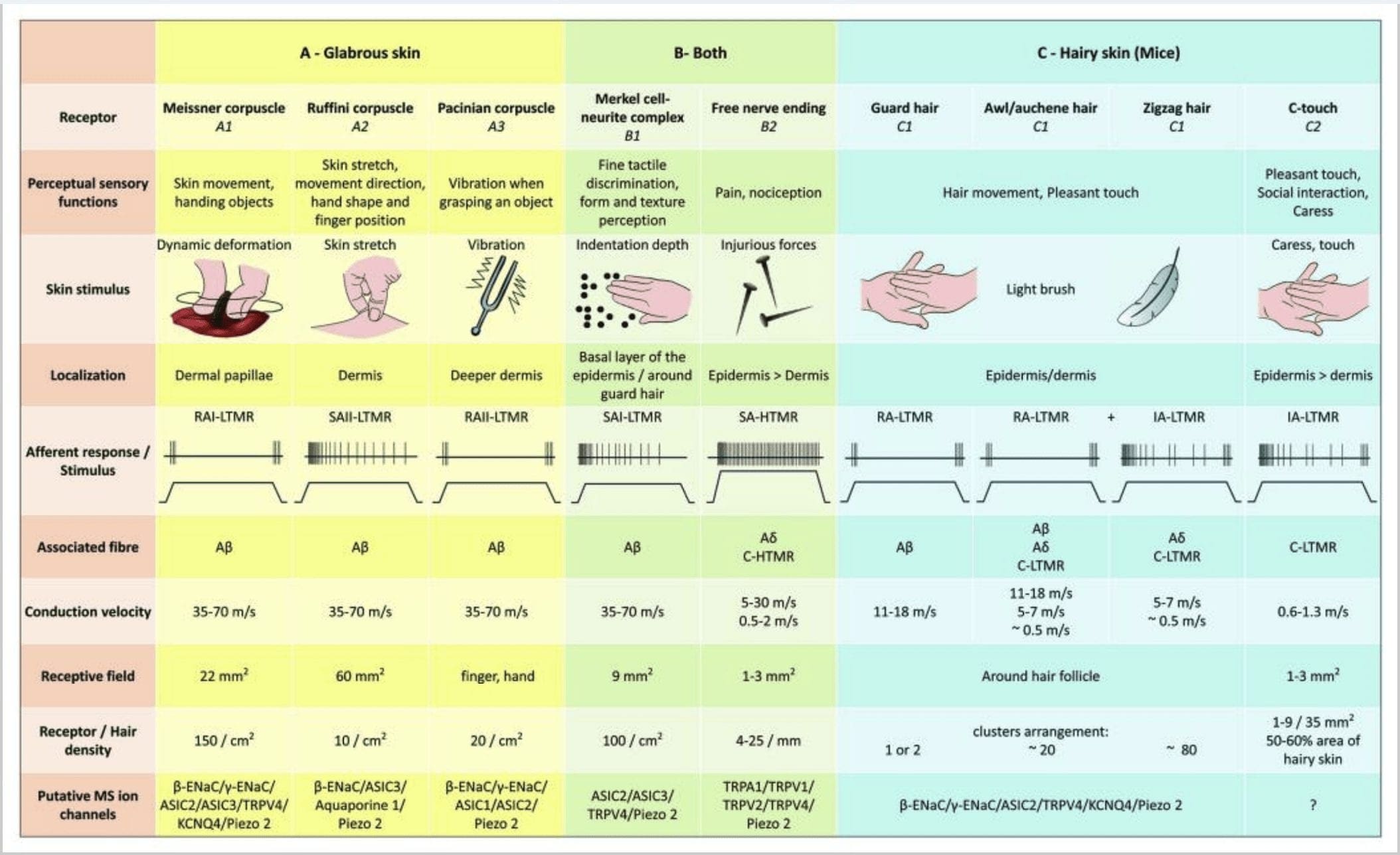
2. Receptorên taktîl ên di mammalan de: Receptorên taktîl ên çerm ji hev cuda dibin têkila bêguneh a ku ji hêla pir receptorên xwedan berbenda mekanîkî ya kêm (LTMRs) di çermê gewre û porî de û têkila zerar a ku ji hêla receptora bendeya mekanîkî ya bilind (HTMRs) ve tê piştgirî kirin, têne cûda kirin. Ew endikên azad ên nervê çêdikin ku bi piranî di epidermisê de bi dawî dibin. (A) Çermê zirav. A1: Laşên Meissner tevgera çerm û şemitîna tiştê di dest de tespît dikin. Ew ji bo radestkirina tiştan û jêhatîbûnê girîng in. Receptor zû bi stimulusê re adapte dibin, bi A? fibers û kêm bi fîberên C û qada receptorên mezin hene. A2: Kulîlkên Ruffini dirêjbûna çerm tespît dikin û ji bo tespîtkirina pozîsyona tiliyê û tiştên destanê girîng in. Receptor hêdî hêdî bi stimulusê re adapte dibe û heya ku stimulus hate sepandin çalakiya xwe didomîne. Receptor bi A ve girêdayî ne? fiber û xwedan qada wergirtina mezin e. A3: Germên Pacinian di dermisê de kûrtir in û vibrasyonê tespît dikin. Receptor bi A ve girêdayî ne? fibers; ew zû bi stimulus re adapte dibin û xwedan qada herî mezin a wergirtinê ne. (B) Tevahiya çerm. B1: Kompleksên Merkel-hucreyê hem di çermê zelal û hem jî li dora porê de hene. Ew bi zexmî di destan de têne diyar kirin û ji bo têgihîştina tevnvîsê û cihêkariya herî baş a di navbera du xalan de girîng in. Ew ji rastbûna tiliyê berpirsiyar in. Receptor bi A ve girêdayî ne? fibers; ew hêdî hêdî xwe bi stimulusê re adapte dikin û xwediyê qada wergir a kurt in. B2: HTMR-yên tûj ên tûj ên bi adaptasyona pir hêdî bi stimulusê re, ango çalak in heya ku stimulasyona nociceptive were sepandin. Ew bi dawiya nerva azad a A-yê têne çêkirin? û C-fîberên bi keratinocytes ve girêdayî ne. (C) Çermê por. C1: Kulîlkên por bi cûreyên cûda yên porê ve girêdayî ne. Di mişkan de mûyên cerdevanan yên dirêjtir û kêm diyarkirî ne, awî/auchenne mezinahiya wan navîn in û zigzag jî porê herî piçûk û herî qels e. Ew bi A ve girêdayî ne? fiber lê ji bo A? û fiberên C-LTMRs ji bo porê awl/auchenne û zizag. Ew di dema lênêrînê de tevgera porê di nav de têkiliyek xweş jî tespît dikin. Ew bi lez an bi kînetîka navîn bi stimulusê re adapte dibin. C2: Endikên nervê yên C-touch bi jêr-cûreyek dawiya fîberên C-yê yên bi dawiya belaş ve ku ji hêla bendek mekanîkî ya nizm ve tête diyar kirin re têkildar in. Pêdivî ye ku ew ji bo hestiyariya xweş a ku ji hêla lênêrînê ve hatî çêkirin kod bikin. Ew bi nermî bi stimulus re adapte dibin û xwedan qada wergirtina kurt in. Kanalên îyonê yên mekanosensîv (MS) yên ku di receptorên cûda yên taktîl de têne diyar kirin li gorî daneyên pêşîn têne destnîşan kirin û hîpoteza heyî ya di bin nirxandinê de kurt dike.
Nifûsa duyemîn a mezin a ku hatî nas kirin endîşeyên A?-LTMR yên di pelikên Awl/Auchenne û zigzag de têkildar e ku bi pelika porê Down re ku bi berfirehî li pisîk û keroşkê hatî lêkolîn kirin were berhev kirin. Ginty û hevkaran destnîşan kirin ku TrkB di astên bilind de di binkomek neuronên DRG-ya navîn-navber de tête diyar kirin. Tomarkirinên hundurîn ên ku bi karanîna ex vivo ya çerm-nervî ya fîberên nîşankirî têne bikar anîn destnîşan kir ku ew taybetmendiyên fîzyolojîkî yên fîberên ku berê li pisîk û kêvroşkê hatine lêkolîn kirin destnîşan dikin: hesasiya mekanîkî ya hêja (Berxa Von Frey < 0.07 mN), zû bersivên li ser teşwîqên tîrêjê vediguhezînin, navîn leza (5.8 ~ 0.9 m/s) û çîpên somayê yên nezivirkirî yên teng.15 Van A?-LTMR dawînên lanceolate yên dirêjî yên ku hema hema bi her pelika porê zigzag û awir/auchene ya qurmê re têkildar in pêk tînin [Fig. 1 (A5)].
Di dawiyê de, wan destnîşan kir ku dawiya dorhêl a bi lez adaptasyona A? LTMR-yên lanceolate yên dirêjî yên ku bi cerdevan (an tylotrich) û pelikên porê awl/auchene ve girêdayî ne, pêk tînin [Hêjî. 1 (A6)].15 Digel vê yekê, porên cerdevanan jî bi kompleksek hucreya Merkel re têkildar in ku qubeyek pêvekirî ya ku bi A? hêdî hêdî adaptasyona LTMR [Hêjî. 1 (A7)].
Bi kurtasî, hema hema hemî pelikên porê zigzag ji hêla herdu dawiya C-LTMR û A?-LTMR lanceolate ve têne nerazî kirin; mûyên awl/auchene sê caran ji hêla A ve têne hundur kirin? Bi lez adapteyî-LTMR, A?-LTMR û C-LTMR dawiya lanceolate; Kulîlkên porê cerdevan ji hêla A? bi lez adapteyî-LTMR dawiya lanceolate dirêj û bi A re têkilî? hêdî hêdî adaptasyona-LTMR-ya dawînên qubeya destanê. Ji ber vê yekê, her pelika porê mişkê hevbendiyên yekta û guhêrbar ên dawiya LTMR-ê ku bi organên dawiya mekanosensory ve ji hêla neurofîzyolojîkî ve cihêreng ve girêdayî ne, werdigire. Dema ku lihevhatina dubare ya van her sê celebên porê dihesibînin, Ginty û hevkarên xwe pêşniyar dikin ku çermê porê ji dubarekirina dubare ya yekîneya derdor pêk tê ku tê de, (1) yek an du mûyên cerdevaniyê yên li navendî, (2) ~ 20 mûyên awl/auchenne yên derdorê û (3) ) ~ 80 mûyên zigzag ên navberkirî [Hêjî. 2 (C1)].
Pêşniyara spinal cord. Pêşniyarên navendî yên A? bi lez adapteyî-LTMRs, A?-LTMR û C-LTMRs bi dawî, lê bi qismî li hev girêdide lamina (II, III, IV) ji stûyê dorsal stûyê spinal. Digel vê yekê, termînalên navendî yên LTMR-yên ku heman pelikên porê an jî cîran di hundurê yekîneyek LTMR ya dorhêl de diherikînin têne hevûdu kirin da ku stûnek LTMR ya teng di qijka piştê ya stûnê de ava bikin [Fig. 1 (B1)]. Ji ber vê yekê, îhtîmalek xuya dike ku kulmek, an stûnek ji endikên hestiyar ên seretayî yên somatotopîkî yên organîzekirî yên di qurmê piştê de lihevhatina pêşnûmeyên navendî yên A?-, A?- û C-LTMR-yên ku heman yekîneya dorhêlê diherikînin û mekanîkî vedigirin temsîl dike. teşwîqên ku li ser heman koma piçûk a pelikên poran tevdigerin. Ginty û hevkarên xwe li ser bingeha jimareya mûyên cerdevan, awer/auchen û zigzag ên qurm û lingan û hejmarên her binkûrek LTMR-ê texmîn dikin ku qurmê pişta mişkê 2,000 £ 4,000 stûnên LTMR dihewîne, ku bi hejmara nêzikî ya peripher re têkildar e. yekîneyên LTMR.15
Digel vê yekê, axonên jêr-tîpên LTMR ji nêz ve bi hevûdu re têkildar in, xwedan pêşnûmeyên pêvekirî û endikên lanceolate yên hevgirtî yên ku heman pelika porê dişoxilînin hene. Wekî din, ji ber ku sê celebên pelika por şikil, mezinahî û pêkhateyên hucreyî yên cihêreng nîşan didin, dibe ku ew xwedan taybetmendiyên birêkûpêkkirina guheztinê an vibrasyonê ne. Van dîtinan bi pîvandinên neurofîzyolojîk ên klasîk ên di pisîk û keroşkê de hevaheng in ku destnîşan dikin ku A? RA-LTMRs û A?-LTMRs dikarin bi guheztina cûrbecûr cûrbecûr pelika porê bi rengek cûda werin çalak kirin.16,17
Di encamnameyê de, têkilkirina di çermê porî de têkelbûna van e: (1) hejmarên têkildar, dabeşkirina cîhê yekta û taybetmendiyên morfolojîk û veguhêz ên cihê yên sê celebên pelikên porê; (2) kombînasyona yekta ya dawiya jêr-tîpa LTMR-ê ku bi her sê celebên porê porê ve girêdayî ye; û (3) hesasiyetên cihêreng, leza guheztinê, şêwazên trêna tîrêjê û taybetmendiyên adaptasyonê yên çar çînên sereke yên LTMR-yên ku bi pelika porê ve girêdayî ne ku rê didin pergala mekanosensory çermê porî ku berhevokên tevlihev ên kalîteyên ku diyar dikin derxe û bigihîne CNS. pêbûn.
Free-Nerve Endings LTMRs
Bi gelemperî, dawiya belaş fîberên C-ê di çerm de HTMRs in, lê binekomek fîberên C-ê bersivê nadin pêwendiya zirardar. Ev jêrkoma aferentên fîbera C-ya takîl (CT) celebek cihêreng ên yekîneyên mekanorceptive yên bêmielînkirî, kêm-sînorê ku di çermê porî, lê ne zelal a mirov û memikan de hene, temsîl dike [Fig. 1 (A8).
Aferentên CT bersivê didin hêzên dakêşanê yên di rêza 0.3-2.5 mN de û bi vî rengî ji deformasyona çerm bi qasî A? afferents.19 Taybetmendiyên adaptasyonê yên aferentên CT bi vî rengî li gorî mekanoreceptorên myelinated hêdî û bilez têne adaptekirin navîn in. Zeviyên wergir ên aferentên CT-ê yên mirovî bi şeklê dordor an ovalî ne. Zeviyek ji yek û neh deqên bersivdar ên piçûk pêk tê ku li ser deverek heya 35 mm2.22 têne belav kirin.50 Receptorên homolog ên mişkî bi şêwazek pişkên domdar hatine organîze kirin ku bi qasî 60-2% ji devera di çermê porê de vedihewîne [Fig. 2 (C23)].XNUMX
Delîlên ji nexweşên ku nebûna aferentên taktil ên myelinated destnîşan dikin ku îşaretkirina di fiberên CT de korteksa însulî çalak dike. Ji ber ku ev pergal di şîfrekirina aliyên cûdakar ên destikê de xizan e, lê ji bo şîfrekirina pêwendiya hêdî, nerm xweş e, fîberên CT-ê yên di çermê por de dibe ku beşek pergalek ji bo hilberandina aliyên xweş û têkildarî civakî yên destikê be.24 Dibe ku aktîvkirina fîberê CT jî di astengkirina êşê de xwedî rolek in û di van demên dawî de hate pêşniyar kirin ku iltîhaba an trawma dikare hestiyariya ku ji hêla LTMR-ya C-fiber ve hatî ragihandin ji têkiliyek xweş berbi êşê biguhezîne.25,26
Kîjan rê rêça CT-afferents hîn nayê zanîn [Fig. 1 (B2)], lê têketinên taktîkî yên kêm-sînor ji şaneyên projekirina spinothalamic re hatine belge kirin,27 bawerî dide raporên kêmasiyên nazik, berevajî yên vedîtina destikê di nexweşên mirovî de piştî hilweşandina van rêçikan piştî prosedurên kordotomiyê.28
LTMR di Çermê Glabrous de
Kompleksên hucre-neurite Merkel û qubeya destan. Merkel (1875) yekem bû ku danasîna histolojîkî ya komikên şaneyên epidermal ên bi navokên lobulkirî yên mezin, danûstendinek bi têlên nervê yên aferent ên texmînkirî re çêkir. Wî texmîn kir ku ew bi navê Tastzellen (hucreyên taktîl) hesta pêwendiyê diparêzin. Di mirovan de, kompleksên hucre-neurît ên Merkel di deverên hestiyar ên çerm de dewlemend dibin, ew di tebeqeya bingehîn a epîdermisê de di tiliyan, lêv û organên zayendî de têne dîtin. Ew di çermê porê de jî bi tîrêjiya kêmtir hene. Kompleksa hucre-neurite ya Merkel ji şaneyek Merkel pêk tê ku ji nêz ve bi termînalek nervê ya mezinbûyî ya ji A-ya yekbûyî ya mîelînekirî ye? fiber [Hêjîrê. 1 (C1)] (nirxandin li Halata û hevkaran).29 Li aliyê epidermal hucreya Merkel pêvajoyên mîna tilikê ku di navbera keratinocytes cîran de dirêj dibin nîşan dide [Fig. 1 (C2)]. Hucreyên Merkel xaneyên epîdermal ên keratinocyte ne.30,31 Têgîna dome touch ji bo navkirina giraniya mezin a kompleksên hucreya Merkel di çermê porê pisîkê de hate destnîşan kirin. Di qubeyek têkilî de dibe ku heya 150 şaneyên Merkel hebin ku ji yek fîberek A? û di mirovan de ji xeynî fîberên A?-ê, A? û fîberên C jî bi rêkûpêk hebûn.32-34
Teşwîqkirina kompleksên hucre-neurît ên Merkel di bersivên Tîpa I (SA I) yên hêdî-hêdî-hêdî de, ku ji qadên wergir ên xalîkirî yên bi sînorên tûj derdikevin, encam dide. Derxistina spontan tune. Van kompleksan bersivê didin kûrahiya lêdanê ya çerm û xwedan çareseriya mekanê ya herî mezin (0.5 mm) ya mekanoreceptorên çerm in. Ew wêneyek cîhêkî ya rastîn a teşwîqên taktîl vediguhezînin û têne pêşniyar kirin ku bibin berpirsiyar ji cûdahiya şekl û tevniyê [Fig. 2 (B1)]. Mişkên ku ji hucreyên Merkel bêpar in, dema ku ew bi rîskên xwe wiya dikin, nikarin bi lingên xwe rûberên tevnvîskirî tespît bikin.35
Ma hucreya Merkel, neuronê hestî an herdu jî cîhên mekanotransductionê ne, hîn jî mijarek nîqaşê ye. Di mişkan de, rûxandina fototoksîkî ya hucreyên Merkel bersiva SA I ji holê radike.36 Di mêşên ku bi şaneyên Merkel ên bi genetîkî hatine tepisandin, bersiva SA I ya ku di amadekirina ex vivo çerm/nerm de hatî tomar kirin bi tevahî winda bû, û destnîşan kir ku hucreyên Merkel ji bo kodkirina rast a Merkel hewce ne. bersivên receptorê.37 Lêbelê, teşwîqkirina mekanîkî ya hucreyên Merkel ên veqetandî yên di çandê de ji hêla zexta motorê ve herikandinên mekanîkî-derî çê nake. Pêvajoyên mîna tiliya hucreya Merkel dikare bi deformasyona çerm û tevgera hucreya epidermisê re tevbigere, û ev dibe ku gava yekem a veguheztina mekanîkî be. Eşkere ye, şert û mercên ku ji bo lêkolîna mekanî-hesasiyeta hucreyên Merkel hewce ne hêj nehatine damezrandin.
dawiya Ruffini. Dawiyên Ruffini bi A? dawiya nervê. Dawiya Ruffini silindirên tevna girêdanê yên piçûk in ku li ser xêzên kolajenê yên dermalî hatine rêz kirin ku ji hêla yek an sê tîrêjên nervê yên myelinated yên bi pîvana 4 m ve têne peyda kirin. Dibe ku di dermisê de heya sê silindirên meyla cihêreng li hev bicivin da ku yek receptorek çêbikin [Fig. 6 (C1)]. Ji hêla strukturî ve, dawiya Ruffini dişibin organên tendona Golgi. Ew bi berfirehî di dermisê de têne diyar kirin û wekî mekanoreceptorên çermî yên ku hêdî hêdî adapteyî celeb II (SA II) têne nas kirin. Li hember paşxaneya çalakiya nervê ya spontan, dakêşînek birêkûpêk a hêdî-hêdî-hêdî ji hêla teşwîqkirina mekanîkî ya domdar a perpendîkular ve an jî ji hêla dirêjbûna dermal ve bi bandortir tê derxistin. Bersiva SA II ji zeviyên wergir ên mezin ên bi sînorên nezelal derdikeve. Receptorên Ruffini di nav şêwaza dirêjbûna çerm de beşdarî têgihîştina arastekirina tevgera tiştan dibin [Fig. 3 (A2)].
Di mişkan de, bersivên SA I û SA II dikarin di amadekirina ex-vivo nervê-çermê de bi elektrofîzyolojîkî werin veqetandin.40 Nandasena û hevkaran immunolocalization ya aquaporin 1 (AQP1) di dawiya Ruffini ya parodontal a mêşhingivên mişkan de ragihand ku pêşniyar dike ku AQP1 beşdarî parastina hevsengiya osmotîk a diranan a ku ji bo mekanotransductionê hewce ye.41 Dawiya Ruffini ya peryodontal jî kanala îyonê ya mekanosensîv ASIC3.42 diyar dike.
Corpuscles Meissner. Korpuskulên Meissner di papillayên dermal ên çermê şêrîn de, bi giranî di kefên dest û lingên lingan de lê di heman demê de di lêv, ziman, rû, di çîp û di organên zayendî de cih digirin. Ji hêla anatomîkî ve, ew ji dawiya nervê ya pêgirtî pêk tê, kapsul ji şaneyên piştgirî yên xêzkirî yên ku wekî lameleyên horizontî yên ku di tevna girêdanê de hatine bicîh kirin têne çêkirin. Yek fiberek nervê A heye? aferentên ku bi laş ve girêdayî ne [Hêjî. 1 (C4)]. Her deformasyonek laşî ya laşê volleyek potansiyelên çalakiyê derdixe holê ku zû diqede, ango, ew bi lez receptoran adapte dikin. Dema ku teşwîq tê rakirin, laş şeklê xwe ji nû ve distîne û dema ku wiya dike voleyek din a potansiyelên çalakiyê çêdike. Ji ber cîhê wan ê rûkalî di dermisê de, van laşan bi bijartî bersivê didin tevgera çerm, tespîtkirina taktîl a şemitandinê û lerzîn (20-40 Hz). Ew ji çermê dînamîk hesas in - mînakî, di navbera çerm û tiştek ku tê destgirtin [Hêjîrê. 2 (A1)].
Corpuscles Pacinian. Korpuskulên Pacinian mekanoreceptorên kûrtir ên çerm in û mekanoreceptorê çermî yê herî hesas in ku di tevgerê de ye. Ev şaneyên hêşînayî yên mezin (bi dirêjahiya 1 mm) ji lameleyên hevseng ên tevna hevgirêdana fibrous û fibroblastên ku ji hêla hucreyên Schwann-ê yên guhezbar ên xêzkirî ve hatine çêkirin, di dermisê kûr de têne xuyang kirin.43 Li navenda laş, di valahiyek dagirtî de, ku jê re ampûla hundurîn tê gotin. , yek A yekta diqedîne? dawiya nervê ya bêmielînkirî aferent [Hêjî. 1 (C5)]. Li ser rûyê çerm bi navendek bi taybetî hesas re zeviyek wergirek mezin a wan heye. Pêşveçûn û fonksiyona gelek celebên mekanoreceptorên ku bi lez têne adaptekirin di mişkên mutant c-Maf de têne asteng kirin. Bi taybetî jî korpûsûlên Pacinian bi giranî atrofî ne.44
Cenazeyên Pacinian di bersivê de li hember ketina çerm, vekêşana nervê ya bi lez adapteyî II (RA II) ku karibe li dû frekansa bilind a teşwîqên vibrasyonê bişopîne, adaptasyona pir bilez nîşan dide.45 Corpuscle Pacinian afferent bi çalakiya demkî ya di destpêk û veqetandina stimulusê de bersivê didin dakêşana domdar. Ji wan re detektorên lezkirinê jî tê gotin ji ber ku ew dikarin guheztinên di hêza stimulusê de tespît bikin û heke rêjeya guherîna stimulusê were guheztin (wek ku di vibrasyonê de çêdibe), bersiva wan bi vê guherînê re têkildar dibe. Laşên Pacinian guheztinên zexta giran û ya herî zêde lerzîn (150-300 Hz) hîs dikin, ku ew dikarin santîmetreyan jî dûrî tesbît bikin [Fig. 2 (A3)].
Bersiva tonîk di laşê Pacinian a dekapsûlkirî de hate dîtin.46 Wekî din, laşên Pacinian ên saxlem di dema teşwîqên domdar ên dakêşanê de bi çalakiyek domdar bersiv didin, bêyî ku tixûbên mekanîkî an frekansa bersivê biguhezînin dema ku sînyala bi navbeynkariya GABA di navbera glia lamellate û dawiya nervê de tê asteng kirin.47 Bi vî rengî. dibe ku hêmanên ne-neuronî yên laşê Pacinian di parzûna stimulasyona mekanîkî û her weha di modulkirina taybetmendiyên bersivê yên neuronê hestî de xwedî rolên dualî bin.
Pêşniyarên spinal cord. Pêşniyarên A?-LTMRs di mêjûya spî de li du şaxan têne dabeş kirin. Şaxa navendî ya sereke di stûnên piştê de di stûnên piştê yên ipsilateral de berbi asta malzarokê ve diçe [Hêjî. 1 (B3)]. Şaxên duyemîn di qurmê piştê de di lamineyên IV de bi dawî dibin û, wek nimûne, di veguheztina êşê de asteng dikin. Ev dibe ku êşê wekî beşek ji kontrola dergehê kêm bike [Fig. 1 (B4)].48
Di astên malzarokê de, aksonên şaxê sereke di du rêçikan de ji hev vediqetin: rêça navîn fasîkula gracil a ku ji nîvê jêrîn ê laş (ling û stûn) agahdarî digihîne pêk tê, û rêça derve jî fasîkula kunîkî ya ku ji nîvê jorîn agahdarî digihîne pêk tê. ya laş (dest û qurm) [Fig. 1 (B5)].
Aferentên taktîl ên seretayî sînapsa xwe ya yekem bi neuronên rêza duyemîn re li medullayê çêdikin ku têlên ji her rêçikê di navokek bi heman navî de dikevin sînapsê: axonên gracile fasciculus di navokê gracil de û aksonên kuçeyî di navokê de sînapse [Fig. 1 (B6)]. Neuronên ku sinapsê werdigirin aferentên duyemîn peyda dikin û tavilê xeta navîn derbas dikin da ku rêyek li aliyê dijberî stûnê mêjî ava bikin - lemniskusa navîn - ya ku di nav stûna mêjî de hilkişiya rawestgeha paşerojê ya di mejiyê navîn de, bi taybetî, di talamusê de [Fig. . 1 (B7)].
Taybetmendiya molekularî ya LTMRs. Mekanîzmayên molekulî yên ku cûrbecûrbûna zû ya LTMR-ê kontrol dikin di van demên dawî de hinekî hatine ronî kirin. Bourane û hevkarên xwe destnîşan kirin ku nifûsên neuronal ên ku receptorê Ret tyrosine kinase (Ret) û hev-receptorê wê GFR?2 di mêşên embryonîk DRG de E11-13 bi bijartî faktora transkrîpsiyonê Mafa îfade dikin.49,50 Van nivîskaran destnîşan dikin ku Mafa/ Neuronên Ret/GFR?2 yên ku di dema zayînê de hatine armanc kirin ku bibin sê celebên taybetî yên LTRMs: neuronên SA1 ku kompleksên hucreya Merkel vedihewînin, noyronên ku zû adapteyî korpusûlên Meissner dikin û aferentên bilez ên ku bi lez adapte dibin (RA I) ku dawiya lanceolate li dora pelikên porê çêdikin. Ginty û hevkar her weha radigihînin ku neuronên DRG-ê ku zû-Ret diyar dikin, bi lez mekanoreceptorên ji laşên Meissner, laşên Pacinian û endikên lanceolate yên li dora pelikên porê vediguhezînin. 51 Ew deverên hedef ên veqetandî di nav navokên dilşewat û cuneate de diherikînin, û şêwazek mekanîzmaya taybetî diyar dikin. projeksiyonên aksonal ên neurone di hundurê mêjî de.
Lêgerîna mekanorceptorên çermê mirovan. Teknîka "microneurografiyê" ku ji hêla Hagbarth û Vallbo ve di sala 1968-an de hatî şirove kirin, ji bo lêkolîna reftarên dakêşanê yên yekta dawînên mekanosensîv ên mirovî yên ku masûlk, movik û çerm peyda dikin, hatîye sepandin (ji bo vekolînê binêre Macefield, 2005).52,53 Piraniya mîkroneurografiya çermê mirovan. lêkolînan fîzyolojiya aferentên taktîl ên di çermê şêrîn a destan de diyar kirine. Qeydên mîkroelektrodê yên ji nervên navîn û ulnar di mijarên mirovan de hestiyariya destikê ya ku ji hêla çar çînên LTMR-an ve hatî çêkirin eşkere kir: Aferentên Meissner bi taybetî ji lêdana ronahiyê li ser çerm hesas in, bersivê didin hêzên rijandina herêmî û leyizkên destpêkî an eşkere yên di qada wergir de. Aferentên Pacinian ji veguheztinên mekanîkî yên bilez re zehf hesas in. Afferent bi tundî bersivê didin ku li ser zeviya wergir difûrin. Kulîlkek Pacinian ku di jimareyek de cih digire dê bi gelemperî bersivê bide lêdana maseya ku destek piştgirî dike. Aferentên Merkel bi taybetî xwedan hestiyariyek dînamîkî ya bilind a li hember teşwîqên xêzkirinê yên ku li deverek veqetandî têne sepandin û bi gelemperî di dema berdanê de bi dakêşanê bersiv didin. Her çend aferentên Ruffini bersivê didin hêzên ku bi gelemperî li çerm têne sepandin, taybetmendiyek bêhempa ya aferentên SA II kapasîteya wan e ku di heman demê de bersivê bidin dirêjbûna çermê laş. Di dawiyê de, yekîneyên porê di pêşiyê de xwedan zeviyên wergir ên ovoid an nerêkûpêk in ku ji gelek deqên hesas ên ku bi mûyên kesane re têkildar in (her qasê ~20 mûyan) pêk tê.
Hestiyariya Mekanîkî ya Keratinocytes
Pêdivî ye ku her teşwîqek mekanîkî ya li ser çerm bi keratinocytes ku epidermis pêk tîne ve were veguheztin. Dibe ku ev hucreyên berbelav ji bilî rolên xwe yên piştgirî an parastinê fonksiyonên nîşankirinê jî pêk bînin. Mînakî, keratinocytes ATP-ê, molekulek girîng a îşaretkirina hestiyariyê, di bersivê de bertekên mekanîkî û osmotîk derdixînin.54,55 Serbestberdana ATP bi stimulasyona otokrîn a receptorên purinergîk dibe sedema zêdebûna kalsiyûmê ya hundurîn. Rêya nîşana kinazê û dûv re avakirina fîbera stresê ya F-aktîn destnîşan dike ku deformasyona mekanîkî ya keratinocytes dibe ku bi rengek mekanîkî destwerdanê li şaneyên cîran bike, wekî hucreyên Merkel ji bo têkiliyek bêguneh û dawiya C-fibera belaş ji bo têkiliyek zirardar [Fig. 55 (C1)].6
Noxious Touch
Mekanoreceptorên sînorê bilind (HTMR) C- û A yên epîdermal in? belaş nerves-endings. Ew bi strukturên pispor ve girêdayî ne û di her du çermê porê de têne dîtin [Fig. 1 (A9)] û çermê zirav [Hêjî. 1 (C7)]. Lêbelê, termê dawiya nervê ya belaş divê bi hişyarî were hesibandin ji ber ku dawiya nervê her gav bi keratinocyte an hucreya Langherans an melanocytes re têkildar e. Analîzên ultrastruktural ên endikên nervê hebûna retîkûlûma endoplazmîkî ya hişk, mîtokondrîya zêde û vezîkula bingehîn a çîn eşkere dike. Parzûnên cîran ên şaneyên epidermal qalind in û di nav tevnên nervê de dişibin membrana post-synaptîk. Têbînî ku danûstendinên di navbera endikên nervê û hucreyên epidermal de dibe ku dualî bin ji ber ku şaneyên epidermal dibe ku navbeynkarên wekî ATP, interleukine (IL6, IL10) û bradykinin serbest berdin û berevajî dawiya nervê peptidergîk dikarin peptîdên wekî CGRP an maddeya P ku li ser hucreyên epidermal tevdigerin serbest berdin. HTMR ji mechano-nociceptors hene ku tenê ji hêla stimulasyonên mekanîkî yên zirardar û nociceptorên polîmodal ve têne heyecan kirin ku di heman demê de bersivê didin germa xeternak û kîmyewî ya biyanî [Fig. 2 (B2)].58
Têlên aferent HTMR li ser neuronên projeksiyonê yên li horna piştê ya stûyê spî bi dawî dibin. A?-HTMR bi neuronên rêza duyemîn bi giranî di lamîna I û V de têkilî daynin, lê C-HTMR di lamîna II de bi dawî dibin [Wêne. 1 (B8)]. Neronên nosîceptîf ên rêza duyemîn ber bi aliyê kontrolê yê stûna spî ve diçin û di maddeya spî de derdikevin, pergala anterolateral ava dikin. Van neuronan bi giranî di talamusê de bi dawî dibin [Fig. 1 (B9 û B10)].
Mechano-Currents Di Neurones Somatosensory
Mekanîzmayên adaptasyona hêdî an bilez a mekanoreceptoran hîn nehatine eşkere kirin. Ne diyar e ka ta çi radeyê adaptasyona mekanoreceptor ji hêla hawîrdora hucreyî ya dawiya nerva hestî, taybetmendiyên xwerû yên kanalên bi mekanîkî-derî ve û taybetmendiyên kanalên îyonê yên voltaja-deriyê axonal ên di neuronên hestî de tê peyda kirin (Wêne. 2). Lêbelê, pêşkeftina vê dawîyê di karakterîzekirina herikên mekanîkî-derî de destnîşan kir ku çînên cihêreng ên kanalên mekanîzmayî di neronên DRG de hene û dibe ku hin aliyên adaptasyona mekanoreceptoran rave bikin.
Tomarkirina in vitro ya di rodiyan de destnîşan kir ku somaya neuronên DRG bi xwezayê mekanosensîv e û herikên mekano-dergehî yên kationîk diyar dike.59-64 Gadolinium û ruthenium sor bi tevahî herekên mekanosensîv asteng dikin, lê kalsiyûm û magnesium ji derve, di giraniya fîzyolojîk de, û her weha amiloride û benzamil, dibe sedema bloka qismî. 60,62,63 FM1-43 wekî astengkerek domdar tevdigere, û derzîlêdana FM1-43 di nav pala paşîn a mişkan de hestiyariya êşê di testa Randall Selitto de kêm dike û bendava vekişîna payê ya ku tê nirxandin zêde dike. bi porên von Frey.65
Di bersivdayîna teşwîqkirina mekanîkî ya domdar de, tîrêjên mekanosensitive bi girtîbûnê kêm dibin. Li ser bingeha domdarên demê yên hilweşîna niha, çar cûreyên cûda yên herikên hestiyar ên mekanîkî hatine veqetandin: tîrêjên bilez ên adapteyî (~ 3~6 ms), tîrêjên navbeynkar (~ 15~30 ms), herikên hêdî hêdî (~200~300 ms). ) û herikandinên pir hêdî hêdî (~ 1000 ms).64 Van herikîn hemî bi bûyera guhêzbar di neronên DRG yên mişkî de hene ku çermê şêrîn ê pişta piştê vedişêrin.64
Hesasiya mekanîkî ya herikên mekanîzmayî dikare bi sepandina rêzek teşwîqên mekanîkî yên zêde were destnîşankirin, ku rê dide analîza stimulus-hera bi hûrgulî.66 Têkiliya herika stimulus bi gelemperî sigmoidal e, û mezinahiya herî zêde ya herikê ji hêla hejmarê ve tê destnîşankirin. kanalên ku di heman demê de vekirî ne.64,67 Balkêş e, ku herikîna mekanosensîv ku bi lez adapte dibe, hate ragihandin ku li gorî heyama mekanosensîv a ultra-hêdî-hêdî tê adaptekirin, sînorê mekanîkî kêm û xala navîn a nîv-çalakkirinê nîşan dide.63,65
Neronên hestiyar ên bi fenotîpên ne-derzbar bi tercîhî herikên mekanosensîv ên bilez ên adapteyî yên bi tixûba mekanîkî ya jêrîn vedibêjin.60,61,63,64,68 Berovajî vê yekê, herikên mekanosensîv ên hêdî û hêdî hêdî têne adaptekirin carinan di şaneyên ne-hezdar64,68 de têne ragihandin. Vê yekê pêşniyar kir ku dibe ku van herikîn beşdarî tixûbên mekanîkî yên cihêreng ên ku di LTMR û HTMR de di vivo de têne dîtin, bikin. Her çend ev ceribandinên in vitro divê bi hişyarî bêne girtin, piştgirî ji bo hebûna di soma neuronên DRG-ê yên mekanotransducersên kêm û berbilind de jî ji hêla teşwîqkirina radîkal-based dirêjbûna nerûnên hestiyar ên mişkî yên çandî ve hate peyda kirin.69 Vê paradîgmayê du eşkere kir. nifûsên sereke yên neuronên hestiyar ên dirêjkirinê, yek ku bersivê dide mezinahiya stimulusê ya nizm û ya din jî ku bi bijartî bersivê dide amplîtuda stimulasyona bilind.
Van encamên girîng, di heman demê de spekulatîf, mekanîzmayî hene: sînorê mekanîkî ya neuronên hestî dibe ku hindik bi rêxistina hucreyî ya mekanoreceptorê re hebe, lê dibe ku di taybetmendiyên kanalên îyonê yên bi mekanîkî-derî de be.
Mekanîzmayên ku di bin hîssazîkirina herikên kationên mekanosensîv ên di neronên DRG yên mişkî de ne di van demên dawî de hatine eşkere kirin.64,67 Ew ji du mekanîzmayên hevdem ên ku bandorê li taybetmendiyên kanalê dikin encam dide: adaptasyon û neçalakkirin. Adaptasyon yekem car di lêkolînên hucreya porê bihîstî de hate ragihandin. Ew dikare ji hêla xebitandinê ve wekî wergerek hêsan a kêşeya aktîvkirina kanala veguherîner a li ser eksê stimulasyona mekanîkî were binav kirin.70-72 Adaptasyon dihêle ku receptorên hestî hesasiyeta xwe ya li hember stimulasyonên nû li ber hebûna stimulusek heyî biparêzin. Lêbelê, perçeyek girîng a herikên mekanosensîv ên di neuronên DRG de li dû teşwîqkirina mekanîkî ya şertî ji nû ve nayê aktîf kirin, ku neçalakkirina hin kanalên veguherîner destnîşan dike.64,67 Ji ber vê yekê, neçalakkirin û adaptasyon bi hev re tevdigerin da ku herikên mekanosensîv birêkûpêk bikin. Van her du mekanîzmayan ji bo hemî herikên mekanosensîv ên ku di neuronên DRG-ya mişkê de têne nas kirin hevpar in, pêşniyar dikin ku hêmanên fizîkî-kîmyayî yên têkildar kînetîka van kanalan diyar dikin.64
Di encamê de, destnîşankirina taybetmendiyên herikên mekanosensîv ên endogenous in vitro di lêgerîna naskirina mekanîzmayên veguheztinê de di asta molekulî de pir girîng e. Cûdahiya ku di bendika mekanîkî de û kinetîka adapteyî ya herikên cihêreng ên dergeh-mekanîkî yên di neuronên DRG de têne dîtin destnîşan dikin ku taybetmendiyên hundurîn ên kanalên îyonê dibe ku, bi kêmî ve beşek, berdêla mekanîkî û kînetîkên adaptasyona mekanoreceptorên ku di dehsalên 1960-an de hatine vegotin rave bikin. 80 amadekariyên ex vivo bikar tînin.
Proteînên Mekanosensîv ên Putative
Herikên îyonê yên mekanosensîv ên di neuronên somatosensory de baş têne destnîşan kirin, berevajî vê, hindik di derbarê nasnameya molekulên ku navbeynkariya mekanotransductionê di mammalan de dikin tê zanîn. Ekranên genetîkî yên li Drosophila û C. elegans, molekulên mekanotransductionê yên berendam, di nav de malbatên kanala Na+ TRP û degenerin/epithelial (Deg/ENaC) jî hene.73 Hewldanên dawîn ên ji bo ronîkirina bingeha molekulî ya mechanotransduction di mammalan de bi giranî li ser van namzedên homologan sekinîne. . Wekî din, gelek ji van berendaman di mekanoreceptorên çerm û neuronên somatosensory de hene (Wêne. 2).
Kanalên Ionê yên Acid-Sensing
ASIC ji binkomek proton-derî ya malbata kanala Na+ ya dejenerîn-epithelial e.74 Sê endamên malbata ASIC (ASIC1, ASIC2 û ASIC3) di mekanoreceptor û nosîceptoran de têne diyar kirin. Rola kanalên ASIC di lêkolînên behrê de bi karanîna mişkan bi jêbirina armanckirî ya genên kanala ASIC ve hatî lêkolîn kirin. Jêbirina ASIC1 fonksiyona mekanoreceptorên çermî naguhezîne lê hesasiyeta mekanîkî ya aferentên ku rûvî diherikînin zêde dike. 75 Mişkên şkestî yên ASIC2 hestiyariyek kêmbûna LTMR-yên çerm ên ku bi lez adapte dibin nîşan didin.76 Lêbelê, lêkolînên paşîn kêmbûna bandorên kASIC2 ragihandin. hem mekano-nerazîbûna visceral û hem jî mekanosensasyona çerm.77 Têkbirina ASIC3 hesasiyeta mekanîkî ya aferentên visceral kêm dike û bersivên HTMR-yên çerm ên li ser teşwîqên zirardar kêm dike.76
Kanala Receptorê Veguhastî
SERmalbata TRP di mammalan de di nav şeş binemalbatan de tê dabeş kirin.78 Hema hema hemî binemalbatên TRP endamên wan ên ku di cûrbecûr pergalên hucreyê de bi mekanosensasyonê ve girêdayî ne.79 Lêbelê, di neuronên hestiyar ên mêjan de, kanalên TRP herî baş ji bo hîskirina agahdariya germî û navbeynkariya iltîhaba neurogenîk têne zanîn. û tenê du kanalên TRP, TRPV4 û TRPA1, di bersivdayîna pêwendiyê de têkildar bûne. Têkbirina îfadeya TRPV4 di mişkan de tenê bandorên hûrgelî li ser bendên mekanosensory akût heye, lê bi tundî hestiyariyê li ser stimulasyonên mekanîkî yên xeternak kêm dike. 80,81 TRPV4 diyarkerek girîng e di şekilkirina bersiva neuronên nociceptive li hember stresa osmotîk û ji hîperalgeziya mekanîkî82,83, di dema iltîhaba de. 1 TRPA1 di hîperalgeziya mekanîkî de rolek xuya dike. Mişkên kêmbûna TRPA1 hestiyariya êşê ya zêde nîşan didin. TRPA84,85 beşdarî veguheztina teşwîqên mekanîkî, sar û kîmyewî di neuronên hestiyar ên nociceptor de dike, lê xuya ye ku ji bo veguheztina hucreya por ne girîng e.XNUMX
Delîlek zelal tune ku destnîşan dike ku kanalên TRP û kanalên ASIC yên ku di mammalan de têne diyar kirin bi mekanîkî têne girtin. Yek ji van kanalên ku bi heterologî têne diyar kirin nîşana elektrîkê ya herikên mekanosensîv ên ku li hawîrdora wana xwemalî têne dîtin vedihewîne. Ev îhtîmala ku kanalên ASIC û TRPs mechanotransducers bin, ji ber nezelaliya gelo kanalek mekanotransduction dikare li derveyî çarçoweya xweya hucreyî bixebite (li beşa SLP3 binêre).
Proteînên Piezo
Proteînên Piezo di van demên dawî de ji hêla Coste û hevkaran ve wekî berendamên sozdar ên ji bo mekanîzmaya proteînan hatine destnîşan kirin.86,87 Vertebrates du endamên Piezo hene, Piezo 1 û Piezo 2, ku berê bi rêzê wekî FAM38A û FAM38B dihatin nasîn, ku di nav eukek pir şaneyek de baş têne parastin. . Piezo 2 di DRG-an de pir e, lê Piezo 1 bi zorê nayê dîtin. Herikandinên mekanosensîv ên piezo ji hêla gadolinium, ruthenium sor û GsMTx4 ve têne asteng kirin (jehrek ji tarantula Grammostola spatulata). Ji Piezo 88. Mîna herikên mekanosensîv ên endogenous, herikên girêdayî Piezo xwedan potansiyelên vegerê yên li dora 1 mV ne û katyonek hilbijartî nîn e, bi Na+, K+, Ca2+ û Mg2+ hemî di kanala bingehîn de derbas dibin. Di heman demê de, herikên girêdayî piezo ji hêla potansiyela membranê ve têne rêve kirin, digel potansiyelên depolarîzekirî de hêdîbûnek berbiçav a kînetîka heyî.1
Proteînên piezo bê guman proteînên mekanosensîner in û gelek taybetmendiyên herikên mekanosensîv ên bilez ên ku di neuronên hestiyar de têne adaptekirin parve dikin. Dermankirina neuronên DRG yên çandeyî bi Piezo 2-ê RNA-ya astengker a kurt, rêjeya neuronên bi herikîna bi lez adapteyî kêm kir û rêjeya noyronên mekanosensîv kêm kir.86 Domên transmembrane li seranserê proteînên piezo hene lê ne motîfên zelal ên ku di nav pore de ne an jî îmzeyên kanala ionê nehatine dîtin. nas kirin. Lêbelê, proteîna mişkê Piezo 1 ku di nav dulaytên lîpîd ên asîmetrîk de hatî safîkirin û vesaz kirin û lîposom kanalên îyonê yên ji rutenyuma sor re hesas in çêdike.87 Di verastkirina mekanotransductionê de bi kanalên Piezo ve gavek bingehîn ev e ku meriv nêzîkatiyên in vivo bikar bîne da ku girîngiya fonksiyonel di îşaretkirina pêwendiyê de diyar bike. Agahdarî di Drosophila de hate dayîn ku jêbirina yek endamê Piezo bersiva mekanîkî ya li ser stimulasyonên zirardar kêm kir, bêyî ku bandorê li pêwendiya normal bike.89 Her çend strukturên wan hîna bêne destnîşankirin, ev malbata nû ya proteînên mekanosensîv mijarek sozdar e ji bo lêkolîna pêşerojê, li derveyî sînor. hestiyariyê. Mînakî, lêkolînek vê dawîyê ya li ser nexweşên bi kêmxwînî (xerocytosis mîrasî) rola Piezo 1 di domandina homeostasis volume ya erythrocyte de nîşan dide.90
Kanala Transmembrane-Mîna (TMC)
Lêkolînek vê dawîyê destnîşan dike ku du proteîn, TMC1 û TMC2, ji bo mekanotransduction şaneya por hewce ne.91 Di mirov û mişkan de ji ber mutasyona gena TMC1 kerrbûna îrsî hat ragihandin. , lê dixuye ku ji bo vekolînê rêberiyek baş e.
Stomatin-Like Protein 3 (SLP3)
Digel kanalên veguheztinê, hin proteînên pêvekirî yên bi kanalê ve girêdayî hatine destnîşan kirin ku di hestiyariya pêwendiyê de rolek dileyzin. SLP3 di neuronên DRG yên mammalan de tête diyar kirin. Lêkolînên ku bi karanîna mişkên mutant ên ku SLP3 tune ne, guhertin di mekanosensasyon û herikên mekanosensîv de destnîşan kirin. 94,95 fonksiyona rastîn a SLP3 nenas dimîne. Wekî ku ji bo C. elegans homolog MEC2.96 Di van demên dawî de GR tê pêşniyar kirin, dibe ku ew di navbera kanala mekanosensîv û mîkrotubulên jêrîn de girêdanek be. Laboratorê Lewin pêşniyar kir ku têlek ji hêla neuronên hestiyar ên DRG ve tê sentez kirin û kanala îyonê ya mekanosensîv bi matrixa derveyî hucreyê ve girêdide.97 Xerabkirina zencîreyê herika RA-mekanosensîv ji holê radike û destnîşan dike ku hin kanalên îyonê tenê dema ku têne girêdan mekanosensîv in. Herikên RA-mekanosensîv jî ji hêla laminîn-332 ve têne asteng kirin, proteînek matrixê ku ji hêla keratinocytes ve hatî hilberandin, hîpoteza modulasyona heyama mekanosensîv ji hêla proteînên derveyî hucreyî ve xurt dike.98
Binmalbata Kanala K+
Bi paralelî herikên mekanosensîv ên depolarîzasyona kationîk, hebûna herikên K+-ya mekanosensîv repolarîzeker di bin lêkolînê de ye. Kanalên K+ yên di şaneyên mekanosensitive de dikarin di hevsengiya heyî de gav bavêjin û beşdarî diyarkirina sînorê mekanîkî û qursa dema adaptasyona mekanoreceptoran bibin.
Endamên KCNK ji malbata kanala K+ ya du-pore (K2P) ne.99,100 K2P rêzeyek rêziknameya berbiçav a ji hêla şaneyên şanayî, laşî û dermankolojîk ve nîşan dide, di nav de guhertinên pH, germahî, dirêjbûn û deformasyona membranê. Van K2P di potansiyela membrana bêhnvedanê de çalak in. Çend binbeşên KCNK di neuronên somatosensory de têne diyar kirin.101 Kanalên KCNK2 (TREK-1), KCNK4 (TRAAK) û TREK-2 di nav çend kanalan de ne ku ji wan re dergehek mekanîkî ya rasterast ji hêla dirêjkirina membranê ve hatî destnîşan kirin.102,103
Mişkên bi gena KCNK2 ya têkçûyî hesasiyeteke zêde ya li hember germê û teşwîqên mekanîkî yên sivik nîşan didin, lê bendeke vekêşanê ya normal ji bo zexta mekanîkî ya zirardar ku bi testa Randall-Selitto li ser piştê tê sepandin, bi karanîna testa Randall Selitto. şert û mercên. Mişkên KCNK104 ji teşwîqkirina mekanîkî ya sivik zêde hesas bûn, û ev hîperhesasiyet bi neçalakkirina zêde ya KCNK2 zêde bû.4 Zêdebûna mekanosensîbilîteya van mêşên nokout dikare were vê wateyê ku dirêjbûn bi gelemperî hem herikên mekanosensîv ên depolarîzekirin û hem jî ji nû vepolarîzekirinê bi rengek hevsengî bi rengek hevsengî çalak dike. depolarîzekirin û repolarîzekirina herikên voltaja-derî.
KCNK18 (TRESK) alîkarek sereke ye ji gihandina paşperdeya K+ ya ku potansiyela parzûna bêhnvedanê ya noyronên somatosensor birêkûpêk dike.106 Her çend nayê zanîn ka KCNK18 rasterast ji teşwîqkirina mekanîkî hesas e, lê dibe ku ew di navbeynkariya bersivên li ser desta sivik de rolek bilîze. her weha teşwîqên mekanîkî yên bi êş. KCNK18 û bi rêjeyek hindiktir KCNK3, têne pêşniyar kirin ku bibin armanca molekularî ya hîdroksî-?-sanshool, pêkhateyek ku di îsota Schezuan de tê dîtin ku receptorên destdanê çalak dike û di mirovan de hestek tirşikê çêdike.107,108
Kanala K+ ya girêdayî voltaja KCNQ4 (Kv7.4) ji bo danîna terciha lez û frekansê ya binpopulasyona mekanoreceptorên ku bi lez adapte dibin hem di mişk û hem jî di mirovan de girîng e. Mutasyona KCNQ4 di destpêkê de bi rengek kerrbûna îrsî ve girêdayî ye. Balkêş e ku lêkolînek vê dawîyê KCNQ4-ê di endikên nervê yên periferîkî yên pelika porê ya ku bi lez adapteyî dike û laşê Meissner de cih digire. Li gorî vê yekê, windakirina fonksiyona KCNQ4 dibe sedema zêdebûnek bijartî ya hesasiya mekanoreceptorê ya li ber vibrasyona kêm-frekansa. Nemaze, mirovên ku ji ber mutasyonên serdest ên gena KCNQ4 windabûna bihîstinê ya dereng dest pê dike, di tespîtkirina lerzînek piçûk, frekansa kêm de performansa pêşkeftî nîşan didin.109

Dr. Alex Jimenez's Insight
Têkilî wekî yek ji hestên herî tevlihev di laşê mirovan de tête hesibandin, nemaze ji ber ku organek taybetî ya ku berpirsiyarê wê tune ye. Di şûna wê de, hesta destgirtinê bi receptorên hestî, ku wekî mekanoreceptor têne zanîn, pêk tê, ku li seranserê çerm têne dîtin û bersivê didin zexta mekanîkî an tehlûkê. Di çermê memikan de çar cureyên sereke yên mekanoreceptoran hene: laşên lamellar, laşên taktîl, endikên nervê yên Merkel û laşên bulboz. Mechanoreceptors ji bo ku destûrê bidin tespîtkirina destikê, ji bo şopandina pozîsyona masûlkeyan, hestî û movikan, ku wekî proprioception tê zanîn, û tewra jî ji bo tespîtkirina deng û tevgera laş kar dikin. Fêmkirina mekanîzmayên avahî û fonksiyona van mekanoreceptoran di karanîna dermankirin û dermankirinên ji bo rêveberiya êşê de hêmanek bingehîn e.
Xelasî
Têkilî hestek tevlihev e ji ber ku ew taybetmendiyên taktîk ên cihêreng, ango lerizîn, şekil, tevnek, kêf û êş, bi performansên cihêreng ên cihêreng temsîl dike. Heya nuha, pevgirêdana di navbera organek têkilî û hesta psîkofizîkî de têkildar bû û nîşangirên molekulî yên çîn-taybet tenê derdikevin holê. Pêşkeftina ceribandinên rodentê ku bi cihêrengiya behreya destikê re têkildar in naha hewce ye ku ji bo hêsankirina nasnameya genomîk a pêşerojê. Bikaranîna mişkên ku kêmasiya binkomên taybetî yên celebên aferentên hestî ne dê nasîna mekanoreceptor û fîberên aferent ên hestî yên ku bi modulek pêwendiyek taybetî ve girêdayî ne pir hêsan bike. Balkêş e, ku gotarek vê dawîyê pirsa girîng a bingeha genetîkî ya taybetmendiyên mekanosensory di mirov de vedike û destnîşan dike ku mutasyonek yek genî dikare bandorek neyînî li hestiyariya destikê bike.110 Ev yek destnîşan dike ku patofîzyolojiya kêmasiya desta mirovan di beşeke mezin de nenas e û bê guman dê Pêşveçûn bi destnîşankirina tam binekoma neuronên hestî yên ku bi modalîteya pêwendiyê an kêmasiyek pêwendiyê ve girêdayî ne.
Di vegerê de, pêşkeftin ji bo danasîna taybetmendiyên biyofizîkî yên herikên mekanî-dergehî pêk hat.64 Pêşkeftina teknîkên nû di van salên dawî de, ku dihêle çavdêriya guheztinên tansiyona parzûnê were şopandin, dema ku herika mekano-derî tê tomarkirin, rêbazek ceribandinê ya hêja ji bo danasînê îsbat kiriye. herikên mekanosensîv ên bi adaptasyona bilez, navber û hêdî (di nav Delmas û hevkaran de hatine nirxandin). LTMR û HTMR.
Xwezaya molekulî ya herikên mekanîzmayî yên di memikan de jî mijarek lêkolînê ya pêşerojê ye. Lêkolîna pêşeroj dê di du perspektîfan de pêş bikeve, yekem ji bo destnîşankirina rola molekula aksesûar a ku kanalên sîtoskeletonê ve girêdide û dê hewce bike ku mekanîzmaya kanalên îyonê yên mîna malbatên TRP û ASIC/EnaC destnîşan bike an birêkûpêk bike. Ya duyemîn, lêkolîna qada mezin û hêvîdar a beşdariya kanalên Piezo bi bersivdana pirsên sereke, têkildarî mekanîzmayên derçûn û dergehê, binekoma neuronên hestî û modalîteyên destikê yên ku Piezo ve girêdayî ne û rola Piezo di hucreyên ne-neronal de têkildar in. mekanosensasyon.
Hestiya destdanê, li gorî ya dîtin, çêj, deng û bîhnê, ku organên taybetî bikar tîne da ku van hestan bişopîne, dikare li seranserê laş bi navgîniya receptorên piçûk ên ku wekî mekanoreceptor têne zanîn pêk were. Cûreyên cihêreng ên mekanoreceptoran dikarin di tebeqeyên cûrbecûr yên çerm de werin dîtin, li wir ew dikarin cûrbecûr teşwîqek mekanîkî bibînin. Gotara li jor hûrguliyên taybetî yên ku pêşkeftina mekanîzmayên avahî û fonksiyonel ên mekanoreceptorên ku bi hestiyariyê re têkildar in destnîşan dike. Agahdariya ku ji Navenda Neteweyî ya Agahdariya Biyoteknolojiyê (NCBI) ve hatî referans kirin. Qada agahdariya me bi chiropractîk û hem jî bi birîn û şert û mercên spinal ve sînorkirî ye. Ji bo nîqaşkirina mijarê, ji kerema xwe ji Dr. Jimenez bipirsin an bi me re têkilî daynin915-850-0900.
Dr. Alex Jimenez

Pirsên din
Êşa piştê yek ji sedemên herî zêde yên seqet û rojên ku di xebata cîhanê de cîhanê winda bûne ye. Ji ber ku rastiyek, êşê paşê wekî duyemîn herî gelemperî ji bo serdana doktorê doktora doktorê, tenê ji hêla infeksiyonên bilind-respirasyonê ve tête kirin. Ji sedî 80 ê nifûsa wê di tevahiya jiyana xwe de bi kêmîrek paşa paşê tecrubeyê dike. Spine avahiyek paqij e ku ji hestî, hevpeyman, ligaments û masûlkan, di nav nermên din ên nerm. Ji ber vê yekê, şert û / an jî mercên aggravated, wekî dîsên herniated, dibe ku dibe ku nîşanên nîşan bide paşê paşê. Girtîgehên werzişî yan birîndarên otobloyê gelek caran ji sedemên paşê paşê paşê têne têne, lê carinan, carinan hêsantirên tevgeran dikarin encamên xweş bibin. Bi dilsoz, alternatîfên alternatîf ên alternatîf, wekî lênêrîna chiropractîk, dikare dikare bi alîkariya karanîna spînal û pisîkên pisîkê, dibe ku bi xilasiya xirabtir dibe baştir bike.

BİXWÎNE BİXWÎNE BİXWÎNE: Rêveberiya Hêza Navnetewî
TEMA ZOREDETIR: Z EXDETAR EX ZDETIR: Pain Pain & Tedawiyên Kronîk
Vala
Çavkanî
Accordion Close
Agahdariya li ser "Mekroksiyonên Structural û Karsazker ên Mirovên Mîkratoran" ne armanc e ku têkiliyek yek-bi-yek bi pisporek lênihêrîna tenduristî ya jêhatî an bijîjkek destûrdar re biguhezîne û ne şîreta bijîjkî ye. Em we teşwîq dikin ku hûn li gorî lêkolîn û hevkariya xwe bi pisporek lênihêrîna tenduristî ya jêhatî re biryarên lênihêrîna tenduristiyê bidin.
Agahdariya Blog & Nîqaşên Berfireh
Qada agahdariya me bi Chiropractic, musculoskeletal, dermanên laşî, tenduristî, beşdariya etiolojîk ve sînorkirî ye nexweşiyên viscerosomatîk di nav pêşandanên klînîkî de, dînamîkên klînîkî yên refleksa somatovisceral ên têkildar, kompleksên subluksasyonê, pirsgirêkên tenduristiyê yên hesas, û / an gotar, mijar û nîqaşên dermanê fonksiyonel.
Em pêşkêş dikin û pêşkêş dikin hevkariya klînîkî bi pisporên ji dîsîplînên cuda. Her pispor ji hêla qada pratîkê ya pîşeyî û dadrêsiya lîsansê ve tê rêve kirin. Em protokolên tenduristî û başbûnê yên fonksiyonel bikar tînin da ku ji bo birîn an nexweşiyên pergala masûlkeyê derman bikin û piştgirî bikin.
Vîdyo, post, mijar, mijar û têgihiştinên me mijarên klînîkî, pirsgirêk û mijarên ku têkildar in û rasterast an nerasterast piştgirî didin qada pratîka meya klînîkî vedigirin.*
Ofîsa me bi awayekî maqûl hewl daye ku navgînên piştgirî peyda bike û lêkolîna lêkolînê an lêkolînên têkildar ên ku postên me piştgirî dikin nas kiriye. Em li ser daxwazê kopiyên lêkolînên piştevaniya lêkolînan ji bo desteyên nîzam û raya giştî peyda dikin.
Em fêhm dikin ku em mijarên ku vegotinek din hewce dike vedihewîne ka ew çawa dikare di nexşeya lênihînê an protokola dermankirinê ya taybetî de bibe alîkar; ji ber vê yekê, ji bo ku em li jor mijara li jor nîqaş bikin, ji kerema xwe bipirsin Dr. Alex Jimenez, DC, An bi me re têkilî bikin 915-850-0900.
Em li vir in ku alîkariya we û malbata we bikin.
Pîroziya
Dr. Alex Jimenez D.C., MSACP, RN*, CCST, IFMCP*, CIFM*, atn*
email: coach@elpasofunctionalmedicine.com
Di nav de wekî Doktorê Chiropractic (DC) destûrdayî ye texas & New Mexico*
Lîsanseya Texas DC # TX5807, New Mexico DC License # NM-DC2182
Wekî hemşîreyek Qeydkirî (RN*) destûrdar e in florida
Florida License RN License de # RN9617241 (Kontrol No. 3558029)
Rewşa kompakt: License de pir-dewletê: Desthilatdar e ku di pratîkê de Dewletên 40*
Alex Jimenez DC, MSACP, RN* CIFM*, IFMCP*, ATN*, CCST Dr.
Karta Karsaziya Min a Dîjîtal





