Tîma Tenduristiya Çîropractîk û Fonksiyonel a Stresa Oksîdatîf a Klînîkê ya Paşê. Stresa oksîdatîf wekî astengiyek di hevsengiya di navbera hilberîna oksîjena reaktîf (radîkalên azad) û parastina antîoksîdan de tê pênase kirin. Bi gotinek din, ew nehevsengiyek di navbera hilberîna radîkalên azad û kapasîteya laş de ye ku bi riya bêbandorkirina ji hêla antîoksîdantan ve bandorên zerardar berteng bike an detoksîf bike. Stresa oksîdatîf di laş de dibe sedema gelek şert û mercên pathophysiolojîk. Di nav wan de nexweşiyên neurodejeneratîf, ango nexweşiya Parkinson, nexweşiya Alzheimer, mutasyonên genê, penceşêr, sendroma westandina kronîk, sendroma Xê ya nazik, nexweşiyên dil û damarên xwînê, atherosclerosis, têkçûna dil, krîza dil, û nexweşiyên înflamatuar hene. Oksîdasyon di çend rewşan de pêk tê:
hucreyên glucose bikar bînin ku enerjiyê bikin
Sîstema parastinê bacteria berbi şer dike û şewitandina avêtinê
cenazeyên qirêj, dermanan, û dûmanê cixarê derxînin
Li gor demên ku di bedenên me de bi milyonan pêvajoyê têne çêkirin ku dibe ku oxygenê dibe. Li vir çend nîşanan hene:
Westînî
Bendava bîranîn û an jî mizgefta bîranînê
Pêçîr û anjî ya hevbeş
Pûçikên bi cilên spî
Kêmbûna çavan
Headaches û hestiyariyê bi dengê
Suspendkirina bi infeksiyonê
Hilbijartina xwarinên organîk û dûrxistina toksînên li hawîrdora we cûdahiyek mezin çêdike. Ev, digel kêmkirina stresê, dikare di kêmkirina oksîdasyonê de bikêr be.
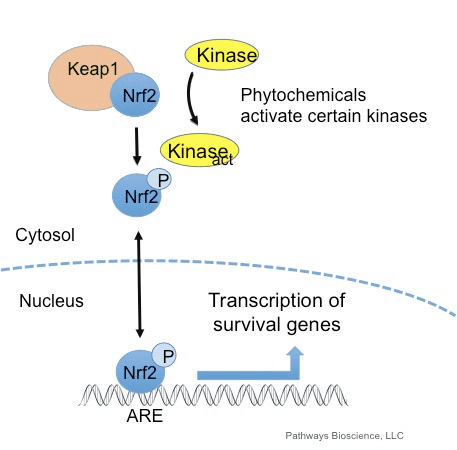
Oksîdantan bi gelemperî di rêbazek kontrolkirî de têne kirin ku ji bo pêvajoyên pêwîst ên di laşê mirovî de, di nav dezgehek, tevlihevkirinê, fonksiyonê parastinê, xweseriyê û bersivên tundûtûjiyê de pêk tê. Lêbelê, hilberên bêgunotkirî yên van oxixantan dikarin beşdar bibin stress, ku dibe ku fonksiyonek hucreyê bandor bike, ji bo pêşveçûnê zextê, nexweşiya kevn û kanser. Mirovanek parastina antioxidant parastina mirovan bi hêla rêgezên giring ên ku bersivên hucreyê bi oxidants kontrol dikin têne rêve kirin. Faktoriya nukleer a erythroid 2-related factor, wekî din jî Nrf2 bi navê tê naskirî ye, rêbazek nûjen dibe ku ji alîyê oxidants re berxwedana hucrekerê hucreyê. Armanca jêrîn jêrê ye ku rola nîqaşkirina Nlf2 di fonksiyona mitochondrial de gotûbêj bikin û nîşan bide.
Abstract
Faktora transkrîpsiyonê NF-E2-girêdayî p45 Faktora 2 (Nrf2; navê genê NFE2L2) di bin şert û mercên stresê de bi rêkûpêkkirina vegotina genê ya torên cihêreng ên proteînên sîtoprotektîf, di nav de enzîmên antîoksîdan, antî-înflamatuar, û detoksîfkirinê jî di nav de rê dide adaptasyon û zindîbûnê. wekî proteînên ku di tamîrkirin an rakirina makromolekulên zirardar de dibin alîkar. Nrf2 di domandina homeostaza redoxa hucreyê de bi rêkûpêkkirina biosentez, bikar anîn, û nûjenkirina glutathione, thioredoxin, û NADPH û bi kontrolkirina hilberîna celebên oksîjena reaktîf ji hêla mitochondria û NADPH oxidase ve rolek girîng heye. Di bin şert û mercên homeostatîk de, Nrf2 bandorê li potansiyela membrana mîtokondrî, oksîdasyona asîda rûn, hebûna substratan (NADH û FADH2 / succinate) ji bo nefesê, û senteza ATP dike. Di bin şert û mercên stresê an teşwîqkirina faktora mezinbûnê de, aktîvkirina Nrf2 li hember zêdebûna hilberîna celebên oksîjena reaktîf ên li mitochondria bi riya veguheztina veguheztinê ya proteîna veqetandî 3 berovajî dike û bi domandina astên faktora respirasyonê ya nukleerî 1 û receptora aktîfkirî ya proliferatorê peroxisome? koaktîvator 1?, û her weha bi pêşvexistina biosenteza nukleotîdê purînê. Aktîvatorên Nrf2 yên dermankolojîk, mîna îsotîocyanate sulforaphane ya xwezayî, vebûna bi navbeynkariya oksîdantê ya pora derbasbûna permeability mîtokondrî û werimîna mîtokondrial asteng dike. Bi balkêş, pêkhateyek sentetîk 1,4-dîfenîl-1,2,3-triazole, ku bi eslê xwe wekî çalakkerek Nrf2 hatî sêwirandin, hate dîtin ku mîtofagiyê pêşve dike, bi vî rengî beşdarî tevestaza mîtokondrîal a giştî dibe. Ji ber vê yekê, Nrf2 di piştgirîkirina yekbûna avahî û fonksiyonel a mitochondria de lîstikvanek girîng e, û ev rol bi taybetî di bin şert û mercên stresê de girîng e.
Keywords:Bioenergetics, Cytoprotection, Keap1, Mitochondria, Nrf2, radîkayên Azad
Highlights
Nrf2 di kampanyona redox homeostasis de parastina rola girîng e.
Nrf2 ê bandora mîkrokê mitokondrial bandor û ATP ê bandor dike.
Nrf2 li oxixationê fatty mitochondrial bandor dike.
Nrf2 yekîtiya avahiyê ya stratejîk û fonksiyona piştgiriya mîkochondrayê dide.
Çalakvanên Nrf2-ê ku bandorên mitochondrial peyman e.
Pêşkêş
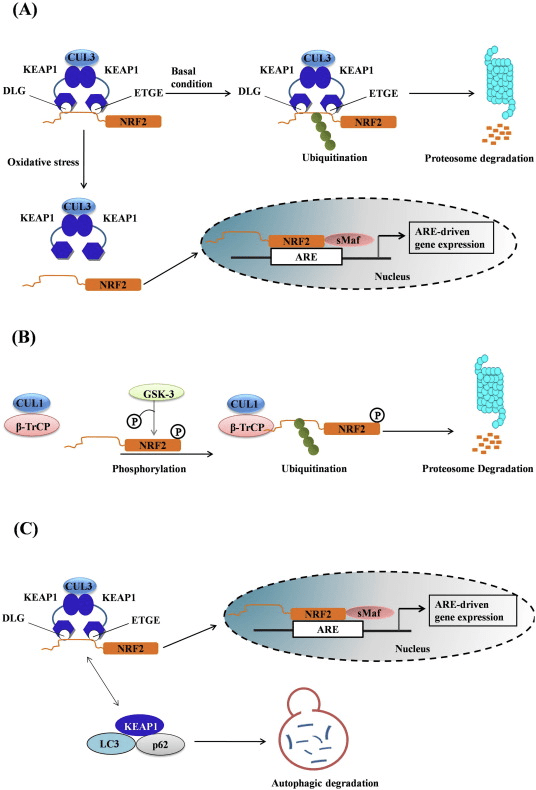
Faktora transkrîpsiyonê NF-E2 faktora 45-ya girêdayî p2 (Nrf2; navê genê NFE2L2) vegotina torên genên ku proteînên bi çalakiyên sîtoprotektîv ên cihêreng kod dikin bi rê ve dibe. Nrf2 bixwe di serî de di asta aramiya proteînê de tê kontrol kirin. Di bin şert û mercên bingehîn de, Nrf2 proteînek demkurt e ku di bin ubiquitination domdar û hilweşandina proteasomal de ye. Sê pergalên ubiquitin ligase yên naskirî hene ku beşdarî hilweşandina Nrf2 dibin. Ji hêla dîrokî ve, yekem regulatorê neyînî yê Nrf2 ku hat keşif kirin proteîna 1-ê ya bi ECH-ê ya mîna Kelch-ê (Keap1) [1] bû, proteînek adapterê substratê ji bo Cullin 3 (Cul3) / Rbx1 ubiquitin ligase [2], [3], [ 4]. Keap1 mekanîzmayek dorhêlî ya pir bikêrhatî bikar tîne da ku Nrf2 ji bo ubiquitination û hilweşandina proteasomal bike armanc, di dema ku Keap1 bi domdarî ji nû ve tê nûve kirin, ku dihêle ku çerx biçe (Hêjî. 1A) [5]. Nrf2 di heman demê de ji hêla glycogen synthase kinase (GSK) 3/?-TrCP-girêdayî Cul1-ligaza ubiquitin [6], [7] ve tê hilweşandin. Herî dawî, hate ragihandin ku, di şert û mercên stresa retîkûlûma endoplazmî de, Nrf2 di pêvajoyek ku bi navbeynkariya E3 ubiquitin lîgaza Hrd1 [8] ve tê ubiquitin û hilweşandin.
1 Nîşaneya Keap1-navîgasyonê ya Nrf2 Ji bo modela bilez a binding and reveneration cyclic. (A) Nrf2 li ser Keap1 dimer bi belaşek bendê ye: yekem bi rêya xwe ya bilind a ETGE (red sticks) dermanên binding û paşê wê ya DLG (laşên reş) yên dakêşî dakêşin. Di vê krîza ya pêvajoyê de, Nrf2 veguherînek nerazî dike û ji bo hilweşîna proteasomal veguherîn e. Free Keap1 nû ve nûvekirin û bi karûbarê Nrf2 ya nû ve tête qedandin, û çiyayê dîsa dîsa dest pê dike. (B) Îroşkerên (diamondên spî) re bi şertên cysteines yên Keap1 (rêkêşên şîn), bi rêvekirina guhertinên adrobatê veguherîn û bêhêz kirin. Keap1 ya nû ve nû ve nûvekirî ye, û neserasyona nû ya Nrf2 agir dike û bi nucleus re wergerandin.
Ji bilî xizmeta proteînek ubiquitin li ser proteîneya substrate veşêre, Keap1 jî ji bo çalakvanên biçûk-molekulên Nrf2 (hilberandinên hilberîn) [9] jî jî jî sensor e. Inducers astengkirina cycle ji rezîlkirina Keap1-navçîtiya ji Nrf2 destê kîmya guherandin bermayiyên cysteine taybetî di nava Keap1 [10], [11] an ji aliyê rasterast Xirabkirina Keap1: Nrf2 virtual qayîm [12], [13]. Ji ber vê yekê, Nrf2 hilweşandiye, û faktoriya veguherînê hilweşîne û wergerandina nucleus (XIG-1B), ku ew bi protein-mafê Mafek Mafek piçûk e, heterodimer dike. bi elementên antîxîdîdant-bersivên biyaniyan, herêmên dezgehên herêmî ên cinsên armancên wê yên bêdeng dikin; û veguherandina destpêkê [14], [15], [16]. Battery ji armancên Nrf2 yên ku bi enzymes yên metabolîzma xenobiotic, protein bi antîxidant û karûbarên antogamatîk, û protokolên proteosomal ên ku di navnîşên xweser ên hûrgelan de rêve dikin û di metroolyona navendî de beşdarî beşdar dikin û tevlî proteînên protein ên cytoprotîkî hene.
Nrf2: Rêveberiya Mamoste ya Cellular Redox Homeostasis
Fonksiyona Nrf2 wekî rêgezek sereke ya homeostasis redox-a hucreyî bi berfirehî tête nas kirin. Vebijandina genê ya her du yekîneyên katalîtîk û birêkûpêk ên ?-glutamyl cysteine ligase, enzîma ku di biosenteza glutathione ya kêmkirî (GSH) de gava gav-sînorker katalîze dike, rasterast ji hêla Nrf2 ve tê rêve kirin [17]. Yekîneya xCT ya pergala xc-, ku sîstenê di nav hucreyan de îtxal dike, di heman demê de hedefek transkrîpsiyona rasterast a Nrf2 ye [18]. Di hucreyê de, cystine vediguhere cysteine, ku pêşengek ji bo biosenteza GSH ye. Digel rola wê di biosenteza GSH de, Nrf2 ji hêla rêziknameya veguheztinê ya hevrêzkirî ya glutathione reductase 1 [19], [20], ku bi karanîna hevrehên kêmkirina NADPH-ê glutathione oksîdkirî ji NADPH re bikar tîne, ji bo domandina glutathione di rewşa wê ya kêmbûyî de amûrek peyda dike. . NADPH-ya pêwîst ji hêla çar enzîmên sereke yên NADPH-çêker ve tê peyda kirin, enzîma malîk 1 (ME1), isocitrate dehydrogenase 1 (IDH1), glukoz-6-fosfat dehydrogenase (G6PD), û 6-phosphogluconate dehydrogenase (PGD), ku hemî ne. ji hêla Nrf2 ve (Hêjî. 2) [21], [22], [23], [24] ve hatiye veguherandin. Bi balkêş, Nrf2 di heman demê de vegotina genê ya înduksîyonî ya formên sîtosolîk, mîkrosomal, û mîtokondrî yên aldehyde dehydrogenase [25], ku NAD(P)+ wekî kofaktor bikar tînin, rê dide NAD(P)H. Bi rastî, astên NADPH û rêjeya NADPH/NADP+ di fîbroblastên embryonîk ên ku ji mêşên Nrf2-knockout (Nrf2-KO) veqetandî ne li gorî şaneyên ji hevpîşeyên wan ên çolê (WT) kêmtir in, û astên NADPH li ser têkbirina Nrf2 li xêzên hucreyên kanserê yên bi Nrf2-ya çalak a bingehîn [26]. Wekî ku tê hêvîkirin, astên GSH di şaneyên ku tê de Nrf2 têkçûyî kêm in; berevajî vê, aktîvkirina Nrf2 bi rêgezên genetîkî an dermankolojîk ve dibe sedema bilindbûna GSH [27], [28], [29]. Ya girîng, Nrf2 di heman demê de vegotina genê ya thioredoxin [30], [31], [32], thioredoxin reductase 1 [28], [29], [32], [33], û sulfiredoksin [34], ku bingehîn in, rê dide. ji bo kêmkirina thiolên proteîna oksîdî.
Nîşan 2 Roja Nrf2 di metabolîzmê ya hucreyên lezgîn ên zûtirîn. Nrf2 hem di milê oksîdatîf [ango, glukoz-6-fosfat dehîdrojenaz (G6PD) û 6-fosfoglukonat dehîdrojenaz (PGD)] û hem jî di milê neoksîdantî de [ango transaldolase 1 (TALDO1) û transketolazê de rêgezek erênî ya genan e ku enzîman kod dikin. TKT)] rêya pentoz fosfatê. G6PD û PGD NADPH çêdike. Nrf2 di heman demê de vegotina genê ya du enzîmên din ên hilberîner ên NADPH, enzîma malîk 1 (ME1) û isocitrate dehydrogenase 1 (IDH1) jî rê dide. Vebijandina genê ya fosforîbosil pyrofosfat amidotransferase (PPAT), ku têketina nav riya biosyntetîk a de novo purine katalîz dike, di heman demê de ji hêla Nrf2 ve jî bi erênî tê rêve kirin, her weha vegotina methylenetetrahydrofolate dehydrogenase 2 (MTHFD2), ku rolek mitondrîkî ya mitondrîkî ye. ji bo biosenteza de novo purine yekîneyên yek-karbonê peyda dike. Pyruvate kinase (PK) bi neyînî ji hêla Nrf2 ve tê rêve kirin û tê çaverê kirin ku ji bo avakirina navberên glycolîtîk û, bi G6PD, metabolîtan re bi riya riya pentose fosfatê û senteza asîdên nukleî, asîdên amînî, û fosfolîpîdan re bibe alîkar. Nrf2 bi neyînî vegotina genê ya ATP-citrate lyase (CL) bi rê ve dibe, ku dibe ku hebûna citrate ji bo karanîna mîtokondrîal an (bi riya issocitrate) ji bo IDH1 zêde bike. Sor û şîn bi rêzê rêziknameya erênî û neyînî nîşan dide. Mîtokondrî bi rengê gewr tê nîşandan. Kurteyên metabolîtan: G-6-P, glîkoz 6-fosfat; F-6-P, fructose 6-phosphate; F-1,6-BP, fructose 1,6-bisphosphate; GA-3-P, glyceraldehyde 3-phosphate; 3-PG, 3-phosphoglycerate; PEP, phosphoenolpyruvate; 6-P-Gl, 6-phosphogluconolactone; 6-PG, 6-phosphogluconate; R-5-P, ribulose 5-fosfat; PRPP, 5-phosphoribosyl-?-1-pyrophosphate; THF, tetrahydrofolate; IMP, inosine monophosphate; AMP, adenosine monophosphate; GMP, guanosine monophosphate.
Roja sereke ya girîng ya Nrf2 wekî rêveberê masterê ya redox homeostasisê ya hucreyê, ew ne ecêb e ku hêjmarên WT-ê, bi riya oxygenên reaksiyonê reaksiyonê (ROS) re di hucreyên ku Nrf2 hate avêtin kirin (Nrf2-KO) mezintir in. [35]. Ev cudahî bi taybetî li ser zehmetiyê bi ajansên ku sedema zextên oksîterê ye. Ji bilî, hucreyên kêm in Nrf2 bi zextên oksîdants yên pir celeb ne hest e û ji hêla Nrf2 inducers, ne, ku di bin şertan de, parastina bandor û demdirêj a ji bo WT cellên WT [29], [36] , [37]. Ji bilî tevahiya tevahiya malpera rûsî ya malostasis, Nrf2 jî ji bo parastina dermankirina redoxê malostasis jî girîng e. Ji ber vê yekê, li gorî WT-ê, tevahiya poolê NADH-NADH-ê ya mitochondrial bi KexXUM-KO di girîng de zêde bûye û di navên Nrf1-KO de kêm dibe kêmkirin [2].
Em bi demek hucreyên zindî bi kar tînin, hêj demên ROS hilberîn di kocultên gloneuronal ên sereke û pişkên mizgefta mizgeftê yên ji WT, Nrf2-KO, yan Keap1-kulê (Keap1-KD) vekirî de çavdêr kirin [38]. Wekî ku hêvî kirin, rêjeya hilberkirina ROSê bi lezgehên Nrf2-KO bi hevalbendên WTyê re bihevtir bûn. Lêbelê, me çavdêriya nerazîkirî ya ku WT bi, Wesayîtên Keap1-KD jî rêjeyên ROSê bilindtir hene, tevî ku heqê cûdahiya di navbera WT û Keap1-KD genotypes de di navbera WT û Nrf2-KO de biçûk bû . paşê em ji ber asta mRNA ji NOX2 û NOX4 analîzê, li subunits bi katalî- ji oxidase du NADPH (nox) isoforms ku li pathology mejî kiryaran de hatine, û dît ku NOX2 ye dramatîk di bin şert û mercên we kêmasîya Nrf2 zêde, lê NOX4 upregulated Gava Nrf2 tevlihevkirî ye, her çend hindek piçûk. Bi rêjeyî, mezinbûna astengkirina di hucrey û tissues ji mûçeyên mutantal de zêdebûna zêdekirina ROS di ROS [38] de. Bi balkêş, ne tenê ne Nrf2 NADPH Oxidase biryara, lê ROS ji hêla NADPH Oxidase ve tête çêkirin dikare dikare Nrf2 çalak bike, wekî hucreyên epîtelial û cardiomyocytes [39], nîşan dide [40]. Di heman demê de, lêkolînek herî nêzî nîşan kir ku NADFNUMX-NADPH-ê çalakiya girêdayî nADPH-NUMX-ê ji bo parastina mirinê ya mirinê û mirinê li ser di zexta zordestiya zronic de [2] ji bo parastina mejmarîzmê girîng e.
Ji bilî çalakiya katalatalî ya NADPH oxidase, respirasyonê ya mitochondular, çavkaniya intracellular yek girîng e ku ROS.By bi karanîna mitokondria-taybetî lêkolînek MitoSOX, me ji bo ROS ya hilberîna mitokondrial ya tevahiya ROS-ê di çarçoveya gloneuronal de kulîlkên sereke yên sereke ji WT, Nrf2-KO, an jî Keap1-KDê [38]. Wekî ku hêvî kirin, hucreyên Nrf2-KO ji hêla Rêjeya ROT ê mitochondrial Rêjeya WTyê bû. Di peymana ku ji bo hilberîna ROS ya tevahî, rêjeya hilberîna ROS-ê ya ROM-ê ya KAP1-KD jî ji hêla WT-cells re dihatin. Girîngî, astengkirina astengiya ku ez bi rotenone re hilberînek di hilberên WT û Keap1-KD de di mitochondrial ROS-ê mitokondrial de, ji hucreyên Nrf2-KO ne bandor bû. Bi berevajî zêdekirina zêdebûna mezinbûna di hilberên ROT-ê ya ROS-ê ya mitotondrial de li ser pyruvate (pisporiya NADH zêde dibe, zêdebûna membrokê mitochondrial, û germkirina normalîn), hilberîna ROS-ê di hucreyên Nrf2-KO kêm kirin. Di hev de, ev peywendiyên berbiçav dikin ku, nebûna Nrf2: (i) çalakiya kampanyona min kêm dibe, (ii) çalakiya tevlihev ya min e ku ji ber sînorên substrates, û (iii) çalakiya zehmet a complex Ez yek ji sedemên sereke ji bo hilberîna mitokondrial ROS e, dibe ku ji ber ku dibe ku ji duyem duyemîn ji elektrîkê veguherîne veguherîne.
Nrf2 Pêkêş û Rizgariya Mitochondrial Membranê bandor dike
Potansiyela membrana mîtokondrî (??m) nîşanek gerdûnî ya tenduristiya mîtokondrî û rewşa metabolê ya şaneyê ye. Di şaneyek saxlem de, ??m ji hêla zincîra nefesê ya mîtokondrî ve tê parastin. Balkêş e, nîşankirinek îzotopîkî ya domdar a bi asîdên amînî di lêkolîna proteomîk a li ser bingehê çandê de di rêza şaneya epîteliya pêsîra mirovî ya MCF10A ya ne-tumorîjenîk a receptor-neyînî ya estrojenê de destnîşan kir ku pêkhateya zincîra veguheztina elektronê ya mîtokondrîal NDUFA4 ji hêla çalakkirina dermankolojîkî ya NDUFA2 ve hatî rêve kirin (ji hêla) di heman demê de verastkirina genetîkî ya Nrf2 (ji hêla Keap1 knockdown ve) rê li ber birêkûpêkkirina binbeşên cytochrome c oxidase COX2 û COX4I1 [42]. Lêkolînek li ser proteoma kezebê bi karanîna elektroforeziya jelê ya du-alî û spektrometriya girseyî ya desorption/iyonîzasyona lazerê ya bi alîkariya matrixê ve hatî dîtin ku Nrf2 vegotina binyekeya synthase ya ATP bi rê ve dibe? [43]. Wekî din, proteîna mîtokondrial DJ-1, ku di domandina çalakiya kompleks I [44] de rolek dilîze, hate ragihandin ku Nrf2 [45], [46] stabîl dike, her çend bandorên neuroprotektîf ên aktîvkirina dermankolojîk an genetîkî yên Nrf2 ji DJ-1 serbixwe ne [47]. Lêbelê, encamên van çavdêriyan ji bo fonksiyona mitochondrial nehatiye lêkolîn kirin.
Li gorî çalakiya astengdar a kompleks I di şert û mercên kêmasiya Nrf2 de, ??m ya bingehîn di fîbroblastên embryonîk ên mişkî yên Nrf2-KO (MEFs) û şaneyên glioneuronal ên seretayî yên çandî de li gorî hevpîşeyên wan ên WT kêmtir e (Hêl. 3, têketin) [35]. Berevajî vê, dema ku Nrf2 ji hêla genetîkî ve bi rêkûpêk ve hatî rêkûpêk kirin (bi qutkirin an qutkirina Keap1) ??m bingehîn bilindtir e. Ev ciyawaziyên ??m di nav genotip de diyar dikin ku nefes ji çalakiya Nrf2 tê bandor kirin. Bi rastî, nirxandina mezaxtina oksîjenê di rewşa bingehîn de eşkere kir ku, li gorî WT, mezaxtina oksîjenê di Nrf2-KO û Keap1-KO MEFs de, bi rêzê ~ 50 û ~ 35%, kêmtir e.
3 Mîrzîzma Pêşniyaz ji bo karê mîtochondrial binçavkirina mercên Nrf2. (1) Asta kêmbûna ME1, IDH1, G6PD, û PGD di asta NADPH-ê de kêm dibe. (2) Asta GSH jî kêm in. (3) Çalakiya kêm a ME1 dibe ku hewza pyruvate ku dikeve nav mitochondria kêm bike. (4) Nifşa NADH hêdîtir e, dibe sedema têkçûna çalakiya kompleks I û zêdebûna hilberîna ROS ya mitochondrial. (5) Kêmkirina FAD bo FADH2 di proteînên mîtokondrî de jî kêm dibe, herikîna elektronê ji FADH2 berbi UbQ û di kompleks III de kêm dike. (6) Damezrandina hêdîtir a UbQH2 dibe ku çalakiya enzîmê ya succinate dehydrogenase kêm bike. (7) Asta zêde ya ROS-ê dibe ku çalakiya kompleksa II-ê bêtir asteng bike. (8) Karbidestiya kêmtir a oksîdasyona asîda rûn dibe sedema kêmbûna hebûna substratê ji bo nefesê mîtokondrî. (9) Glîkolîz wekî mekanîzmayek tezmînatê ji bo kêmbûna hilberîna ATP-ê di fosforîlasyona oksîdatîf de zêde dibe. (10) ATP synthase berevajî tevdigere da ku ??m biparêze. Sor û şîn, bi rêzê, birêkûpêkbûn û dakêşandinê destnîşan dikin. Qutik hebûna delîlên ceribandinê nîşan dide. Di navberê de wêneyên mîtokondrîyên astrosîtên kortikal ên WT û Nrf2-KO yên ku ji hêla sondaya fluorescent a potensiyometrîk tetramethylrhodamine methyl ester (TMRM; 25 nM) ve têne xuyang kirin nîşan dide. Barê pîvanê, 20 �m.
Van ciyawaziyên di ??m û nefesê de di nav genotip de ji hêla rêjeya karanîna substratan ji bo nefesa mîtokondrî ve têne xuyang kirin. Serîlêdana substratan ji bo çerxa asîda tricarboxylic (TCA) (malate/pyruvate, ku di encamê de hilberîna substrata I kompleksa NADH zêde dike) an metil succinate, substratek ji bo kompleksa II, dibe sedema zêdebûna gav bi gav di ??m de di her du WT de. û neuronên Keap1-KD, lê rêjeya zêdebûnê di hucreyên Keap1-KD de zêdetir e. Ya girîngtir, şeklên bersiva van substratên çerxa TCA di navbera her du jenotîp de cihêreng in, bi vî rengî bilindbûna bilez a ??m di hucreyên Keap1-KD de li ser lêvekirina substratê bi daketinek bilez li şûna deştê tê şopandin, ku pêşnîyarek bêhempa dide. vexwarina substratê ya bilez. Van dîtinan bi astên pir kêmtir (bi 50%) yên malate, pyruvate û succinate re ku piştî 70-h pulse [U-1C13] glukozê di Keap6-KO de li gorî WT MEF hatine dîtin re lihevhatî ne. şaneyên [1]. Di neuronên Nrf24-KO de, tenê pîrûvat dikare ??m zêde bike, lê malate û metil suksînate dibe sedema depolarîzasyona sivik. Bandora Nrf2 li ser hilberîna substratê mîtokondrî mekanîzmaya sereke ye ku Nrf2 bandorê li fonksiyona mîtokondrî dike. Indeksa redoksa NADH ya mitokondrî (hevsengiya di navbera vexwarina NADH-ê ji hêla I-ya kompleks û hilberîna NADPH-ê di çerxa TCA de) di şaneyên Nrf2-KO de li gorî hevpîşeyên wan ên WT-ê pir kêmtir e, û ji bilî vê, rêjeyên nûjenkirina hewzên NADH û FADH2 piştî astengkirina kompleksa IV (bi karanîna NaCN) di hucreyên mutant de hêdîtir in.
Di mîtokondrîyên ku ji mêjî û kezeba mûşîyê veqetandî ne, lêzêdekirina substratan ji bo kompleksa I an jî ji bo kompleksa II dema ku Nrf2 tê çalak kirin û bi bandortir gava ku Nrf2 têk diçe rêjeya vexwarina oksîjenê bi hêztir zêde dike [35]. Ji ber vê yekê, malat di Keap1-KD de li gorî WT rêjeyek bilindtir vexwarina oksîjenê çêdike, lê bandora wê di mitochondria Nrf2-KO de qelstir e. Bi heman rengî, di hebûna rotenone de (gava ku kompleks I tê asteng kirin), suksînate li Keap1-KD-ê li gorî WT-ê bi rêjeyek mezintir vexwarina oksîjenê çalak dike, di heman demê de bersiv di mitochondria Nrf2-KO de kêm dibe. Wekî din, çandên neuronal ên seretayî yên Nrf2-KO û mêş ji jehrîbûna înhîbîtorên kompleks II 3-nitropropionic acid û malonate re hesastir in, di heman demê de veguheztina intrastriatal a astrocytes Nrf2-zêde derbirrîn parastin e [48], [49]. Bi heman rengî, mêşên Nrf2-KO ji wan re hesastir in, di heman demê de ku aktîvkirina genetîkî an dermankolojîk a Nrf2 bandorên parastinê li dijî, neurotoksîkiya ku ji hêla 1-methyl-4-phenylpyridinium ion 1-methyl-4-phenylpyridinium ve hatî çêkirin bandorên parastinê hene. Modela heywanan a 1,2,3,6-tetrahydropyridine ya nexweşiya Parkinson [49], [50], [51], [52], [53], [54], [55], [56], [57], [58], [59], [60], [61].
Rêjeya kontrolkirina nefesê (RCR), rêjeya Dewleta 3 (ADP-teşwîqkirî) ji nefesa Dewleta 4 re (ADP tune), di nebûna Nrf2 de kêm dibe, lê RCR di navbera mîtokondrîya Keap1-KD û WT de wekhev e [35 ]. Ji ber ku RCR nîşanek asta hevgirtina çalakiya zincîra respirasyonê ya mîtokondrî ya bi fosforîlasyona oksîdatîf e, ev vedîtin destnîşan dike ku rêjeya bilind a nefesê di mîtokondrîya Keap1-KD de ne ji ber veqetandina fosforîlasyona oksîdatîf e. Zêdetir ew pêşniyar dike ku dema ku Nrf2 tê çalak kirin fosforîlasyona oksîdatîf bikêrtir e. Rêjeya bilind a nefesê di mîtokondrîya Keap1-KD de bi astên bilindtir ên hilberîna ROS-ê yên mîtokondrî [38] re hevaheng e ji ber ku rêjeyên nefesê yên bilind dibe ku bibe sedema zêdebûna lehiya elektronê. Lêbelê, di bin şert û mercên stresa oksîdasyonê de, hilberîna ROS-ê ya zêde ji hêla veguheztina veguheztinê ya girêdayî Nrf2-a proteîna veqetandî 3 (UCP3) ve tê berevajîkirin, ku guheztina protonê ya membrana hundurîn a mitochondrial zêde dike û di encamê de hilberîna superoksîdê kêm dike [62]. Di van demên dawî de, hate destnîşan kirin ku hilbera peroksîda lîpîdê 4-hîdroksî-2-nonenal navbeynkariya bilindkirina Nrf2-girêdayî UCP3 di kardiomyocytes de dike; ev dibe ku bi taybetî ji bo parastinê di bin şert û mercên stresa oksîdasyonê de yên wekî yên di dema reperfuzyonê ya ishemiyayê de girîng be [63].
Nrf2 Affekeriya Xweseriya Oxoxative û Têkiliya ATP
Li gorî bandora Nrf2 li ser nefesê, di mîtokondrîya mêjî û kezebê de, kêmasiya Nrf2 dibe sedema kêmbûna kargêriya fosforîlasyona oksîjenê (wek ku ji hêla rêjeya ADP-ê bi oksîjenê ve, ku ji bo senteza ATP-ê tê vexwarin) tê texmîn kirin, lê çalakkirina Nrf2 (Keap1 -KD) bandora berevajî heye [35]. Li gorî WT-ê, astên ATP di hucreyên bi vesazkirina damezrîner a Nrf2 de pir zêde ye û dema ku Nrf2 tê hilweşandin [64] an têkçûn [35] kêmtir e. Wekî din, karanîna înhîbîtorên fosforîlasyona oksîdatîf (oligomycin) an glycolîzê (îodoacetic acid) eşkere kir ku Nrf2 awayê ku hucreyên ATP hilberîne diguhezîne. Bi vî rengî, di neuronên WT de, oligomycin dibe sedema daketinek bêkêmasî ya ATP û asîda iodoacetic bandorek din tune. Balkêş e, di hucreyên Nrf2-KO de, oligomycin astên ATP zêde dike, ku dûv re hêdî hêdî, lê bi tevahî, ji hêla iodoacetic acid ve tê qelandin, ev destnîşan dike ku di nebûna Nrf2 de, glycolîz, û ne fosforîlasyona oksîtîf, çavkaniya sereke ya hilberîna ATP ye. Balkêş e, tevî ku zêde karîgerîya fosforîlasyona oksîtative di hucreyên Keap1-KD de, lêzêdekirina oligomycin dibe sedema kêmbûna ~ 80% di asta ATP de, û acid iodoacetic dibe sedema kêmbûna ~20% din. Bi vî rengî, an kêmbûna Nrf2 an jî aktîvkirina wê ya damezrîner beşdariya fosforîlasyona oksîdatîf kêm dike û tevkariya glycolîzê berbi senteza ATP-ê zêde dike. Ev bandor bi taybetî dema ku Nrf2 tune be diyar dibe û bi girêdayîbûna ??m bi hebûna glukozê ya di navgîniyê de [35] û astên zêde yên navbeynkarên glîkolîtîk (G-6-P, F-6-P) re hevaheng e. , dihydroxyacetone fosphate, pyruvate, û lactate) piştî hilweşandina Nrf2 [24].
Zêdebûna asta ATP piştî astengkirina F1F0-ATPase ji hêla oligomycin ve destnîşan dike ku di nebûna Nrf2 de, F1F0-ATPase wekî ATPase û ne ATP-sentazek kar dike, ango, ew berevajî tevdigere. Veguheztinek wusa di çalakiyê de bi îhtîmalek mezin hewcedariya pompkirina protonan li seranserê parzûna mîtokondrî ya hundurîn destnîşan dike da ku hewl bidin ku ??m, ya ku ji bo yekbûna fonksiyonel a vê organelê girîng e, biparêze. Berevajîkirina fonksiyona F1F0-ATPase di heman demê de ji hêla depolarîzasyona mîtokondrî ya ku li ser rêveberiya oligomycin li ser hucreyên Nrf2-KO-yê hatî dîtin ve tê xuyang kirin, ku berevajî hîperpolarîzasyona ku di hevpîşeyên wan ên WT an Keap1-kêmasî de çêdibe [35]. Bi tevayî, wusa dixuye ku di bin şert û mercên kêmasiya Nrf2 de ATP di serî de di glîkolîzê de tê hilberandin, û ev ATP paşê ji hêla F1F0-ATPase ve ji bo domandina ??m tê bikar anîn.
Nrf2 Oxuxation Mittyondrial Fatty Acid
Bandora kêmasiya Nrf2 li ser ??m bi taybetî diyar dibe dema ku şaneyên di navgîniya bê glukozê de têne inkub kirin, û ??m di Nrf50-KO de li gorî şaneyên WT ~2% kêmtir e [35]. Di bin şert û mercên kêmbûna glukozê de, oksîdasyona asîda rûnê ya mîtokondrî (FAO) peydakerek sereke ya substratan e ji bo nefes û fosforîlasyona oksîdatîf, ku pêşniyar dike ku Nrf2 dikare bandorê li FAO bike. Bi rastî, karîgeriya FAO hem ji bo asîda rûnê ya palmîtîk a têrkirî ya zincîra dirêj (C16:0) hem jî asîdeya hexanoîk a zincîra kurt (C6:0) di Keap1-KO MEF û mîtokondrîyên dil û kezebê yên veqetandî de ji ya wan zêdetir e. Hevalên WT, di heman demê de ew di hucreyên Nrf2-KO û mitochondria de kêmtir e [65]. Van bandoran ji mirovan re jî pir têkildar in: bi rastî, guhertinên metabolîk ên ku nîşana yekbûna çêtir a FAO-yê bi çalakiya çerxa TCA-yê didin, di lêkolînên destwerdana mirovî de bi parêzên bi glukoraphanîn ên dewlemend, pêşengê aktîvatorê klasîk Nrf2 sulforaphane, hatine ragihandin. 66].
Di gava yekem a FAO ya mîtokondrî de, hîdrojena pro-R ya ?-karbonê wekî hîdrojen derdikeve ku kofaktora FAD-ê ji FADH2 kêm dike, ku di encamê de elektronan vediguhezîne ubiquinone (UbQ) di zincîra nefesê de, di dawiyê de beşdarî hilberîna ATP dibe. . Digel ku teşwîqkirina FAO ji hêla palmitoylcarnitine ve di nebûna glukozê de dibe sedema zêdebûna bendewar a asta ATP di şaneyên WT û Keap1-KO de, digel ku bilindbûna ATP di hucreyên Keap1-KO de zûtir e, dermankirina heman rengî di Nrf2-KO de guheztina ATP-ê çênabe. MEFs [65]. Vê ceribandinê destnîşan dike ku, di nebûna Nrf2 de, FAO tê tepisandin, û wekî din, ew tepisandina FAO wekî yek ji sedemên kêmbûna asta ATP-ê di bin şert û mercên kêmasiya Nrf2 de destnîşan dike [35], [64].
Bi taybetî, mirovên 293-T cellên ku Nrf2 hatiye rawestandin heye ku gotina CPT1 û CPT2 [67], pisporên palmitoyltransferase (CPT), rêjeya rêjeya enzyme ya FAO ya mitochondrial. Di peymanê de, Rêzên mRNA yên Cpt1 di binavên NTf2-KO de bi karûbarên WT-ê re kêm in. [68]. CPT ji bo veguhastina komela acylê ya acyl-CoA ji coenzyme A ji l-carnîtine veguhastin û bi vî awayî veguherîna acylcarnitine ji cytoplasmê di mitochondrayê de destûrê dide. Tevî ku ev nehatiye tehlîmkirin, lê dibe ku ji bilî bandora CPT1, Nrf2 dikare karanîna enzyme bandor bike ku bi asta niştecîhên sereke yên bêdengî, malonyl-CoA jî kontrol bikin. Ji ber vê yekê, ji aliyê mekanîzmîzma ku ne diyar e, Nrf2 veguhastina têgotina CoA-desaturase (SCD) [69] û lyase citrate (CL) [69], [70]. Hişyarî, kişandin an anîkirina SCD ya SCD dibe sedema fosphorylasyon û çalakiya AMP-çalak protein kinase (AMPK) [71], [72], [73], û dibe ku ev nirxandin ku, di nehf2 de, asta SCD dê di zêdekirina çalakiya AMPK de zêde bibe. Ev dikare ji hêla hilberîna protein ya AMPK ve tê dîtin ku di navendên Nrf2-KO de hate dîtin [68], dîtina ku peymanek nêzîkî bi zêdebûna AMPK ve ye, ku di navendên Keap1-KD de hatine ragihandin. mice [74]. Di encamê de encama çalakiya AMPK ya ku ji hêla Nrf79 ve çalakiya nrf75 ve tê veşartî ye, ji bo fosphorylation (li Ser2) ya acetyl-CoA carboxylase (ACC) [2] ye, dibe ku bêtir veguherî li ser Nrf70 bêtir veguherandin. [2 ]. Çalakiya ACC-a, di tevlîheviya CL-ê de ku dê hilberîna acetyl-CoA zêde bike, ji bo sektora ACC, dikare dibe ku hilberên ACC-ê, malonyl-CoA zêde bike. Bila asta malonyl-CoA dê CPT bike, bi vî awayî veguhastina acîdê fattyê di mitochondria. Di dawiyê de, Nrf36 bi awayekî erênî bişkojka CD76 [2], wergera ku wergera pisporê li ser plasmma û membranên mitochondrial veguherîne. Ji ber vê yekê, yek mekanîzmîzma ku NrfXNUMX dikare bandoriya bandora FAO ya mitochondrial bandor dike ku bi veguhastina veguherîna dirêjiya zincê zêrîn ya mîkochondrayê ye.
Ji bilî rêbazê rasterast ya rasterast-nivîskî, Nrf2 jî dikare bandorkirina karûbarên mitokondrial FAO bi bandorên metabolîzmê re li ser sekultera hucreyê. Ev dikare bi taybetî dema ku çalakiya Nrf2 kêm kêm an ne, ne rewş e ku rewşa dewletê ya oksîdekirê veguherîna hucreyê ya hucreyê veguherîne. Bêguman, gelek enzîmên FAO têne guhartin ku guhartinên redoxî têne naskirin. Yek ji van enzyme de acyl-ayd dehydrogenase (VLCAD) pir pir zêrîn e, ku ji 80% zêdetir çalakiya palmitoyl-CoA dehydrogenasyon di nav destên mirovan de [77] dike. Balkêş, Hurd et al. [78] diyar kir ku VLCAD di nav deverên cysteine de hene ku bi guhertinên xwe veguherîne li ser rûyê rûyê dil mitochondria bi H2O2 re biguherînin. Additionally, S-nîtrosîlasyonê ya VLCX li Sys238 li ser enerjiya katalîtîk [79] çêtir dike, û dibe ku oksîdasyonê ya heman cysteine dibe ku bandoreke dijberî, paşê bi bandorkirina FAO mitochondrial. Ji ber vê yekê ew gengaz e ku, tevî Vladîpêşên Rêjeya VLCAD ne li WT, Nrf2-KO, yan Keap1-KO Mêrs [65], çalakiya enzyme ya VLCAD dikare di asta Nrf2 de ji ber asta bilind ROS
Li ser bingeha van hemî dîtinan, dikare were pêşniyar kirin ku (Wêne. 3): Di nebûna Nrf2 de, astên NADPH kêmtir in ji ber kêmbûna îfadeya ME1, IDH1, G6PD, û PGD. Asta kêmbûna glutathione di heman demê de ji ber kêmbûna îfadeya enzîmên ku beşdarî biosentez û nûjenkirina wê dibin û astên jêrîn ên NADPH-ê yên ku ji bo veguheztina oksîdankirî ji forma kêmkirî ya glutathione re hewce ne kêmtir in. Vebijandina kêm a ME1 dê hewza pîrûvatê ya ku dikeve nav mitochondria kêm bike, digel ku glycolîz dibe çavkaniya sereke ya piruvatê. Nifşa NADH hêdîtir e, ku dibe sedema têkçûna çalakiya kompleks I û zêdebûna hilberîna ROS ya mitochondrial. Kêmkirina FAD bo FADH2 di heman demê de hêdîtir e, bi kêmî ve ji ber oksîdasyona asîda rûnê kêmtir bikêrhatî ye, ku herikîna elektronê ji FADH2 berbi UbQ û berbi kompleks III xeternak dike. Ji ber ku UbQH2 çalakkerek succinate dehydrogenase ye [80], hêdîbûna avakirina wê dibe ku çalakiya enzîmê ya succinate dehydrogenase kêm bike. Asta zêde ya superoxide û hîdrojen peroksîtê dikare çalakiya kompleks II bêtir asteng bike [81]. Karbidestiya kêmtir a oksîdasyona asîda rûn dibe sedema kêmbûna hebûna substratê ji bo nefesê mîtokondrîal û hilberîna ATP di fosforîlasyona oksîdatîf de. Wekî mekanîzmayek tezmînatê, glycolîz zêde dibe. ATP synthase berevajî, wekî ATPase, di hewildana domandina ??m de kar dike.
Nrf2 û Biogenesis Mîtochondrial
Hat ragihandin ku, li gorî WT, kezeba mişkên Nrf2-KO xwedî naverokek mîtokondrî ya kêmtir e (wek ku ji hêla rêjeya mîtokondrial bi DNAya nukleer ve hatî destnîşankirin); ev di hem mêşên WT û hem jî Nrf24-KO de ji hêla rojiya 2-h ve bêtir kêm dibe; berevajî vê, her çend di bin şert û mercên xwarina normal de ji WT cûda nebe jî, naveroka mîtokondrî ya di mişkên bi çalakiya Nrf2 ya bilind de ji rojiyê bandor nabe [82]. Balkêş e, lêzêdekirina bi aktîvatorê Nrf2 (R)-?-lipoic acid [83], [84], [85] di adipocytes 3T3-L1 de biyojeneya mitokondrîal pêşve dike [86]. Du çînên rêgezên veguheztina navokî di biyojeneya mitokondrî de rolek girîng dileyzin. Çîna yekem faktorên veguheztinê ne, wek faktorên respirasyonê yên nukleer11 û 2, ku îfadeya genên ku binbeşên pênc kompleksên respirasyonê kod dikin, hêmanên werger ên mîtokondrî, û enzîmên biosyntetîk ên heme yên ku li matrixê mîtokondrîal têne veguheztin kontrol dikin [88]. Piantadosi et al. [89] destnîşan kir ku verastkirina transkrîpsiyonê ya girêdayî Nrf2 ya faktora respirasyonê ya nukleerî 1 biyojeneya mitokondrîal pêşve dike û li dijî cytotoxicitiya kardiotoksîkî ya kemoterapî anthracycline doxorubicin diparêze. Berevajî vê, Zhang et al. [82] ragihandine ku aktîvkirina genetîkî ya Nrf2 bandorê li îfadeya mRNA ya bingehîn a faktora respirasyonê ya nukleerî 1 di kezeba mûşî de nake.
Çîna duyemîn a rêgezên veguheztina navokî yên bi fonksiyonên krîtîk ên di biyojeneya mîtokondrî de hevaktîvatorên transkrîpsyonel in, wek receptorê aktîfkirî yê proliferatorê peroxisome? hevaktîvator (PGC)1? û 1?, ku bi faktorên veguheztinê, makîneya veguheztina bingehîn û ARN-hevberdanê, û enzîmên guhêrbar-histone [88], [90], [91] re têkildar in. Daxuyaniya malbata PGC1 ya hevaktîvator ji hêla gelek nîşanên hawîrdorê ve tê bandor kirin. Dermankirina fîbroblastên mirovî bi aktîvatorê Nrf2 sulforaphane dibe sedema zêdebûna girseya mîtokondrîal û vegirtina PGC1? û PGC1? [92], her çend pêwendiya potansiyel a li ser Nrf2 di vê lêkolînê de nehat lêkolîn kirin. Lêbelê, mişkên bi diyabetî yên ku tê de Nrf2 an ji hêla gena Keap1 ve tê çalak kirin (db/db:Keap1flox/?:Nrf2+/+) an jî têkçûn (db/db:Keap1flox/?:Nrf2?/?) xwedî PGC1 kezebê kêmtir in? astên derbirînê ji heywanên kontrolê (db/db:Keap1flox/+:Nrf2+/+) [93]. Di asta mRNA de ji bo PGC1 cudahî tune? di kezeba mişkên ne-diyabetî de ku WT an Nrf2-KO ne de têne dîtin, di heman demê de ev ast di heywanên Nrf2-zêde îfade (Keap1-KD û Keap1-KO-taybetî yên kezebê) de kêmtir in [82]. Nemaze, rojiyek 24 demjimêran astên PGC1 zêde dike? mRNA di kezeba mişkên hemî genotip de, lê zêdebûn di kezeba Nrf2-KO de li gorî mêşên WT an Nrf2-zêde derbirrîn bi girîngî mezintir e. Li gorî WT, mêşên Nrf2-KO yên ku ji ber enfeksiyonê bi enfeksiyona septîk an birîna pişikê ya akût re rûbirû ne, verastkirina transkripsiyonê ya faktora 1 û PGC1 ya respirasyonê ya nukleerî nîşan dide? [94], [95]. Bi hev re, van çavdêriyan pêşniyar dikin ku rola Nrf2 di domandina astên her du faktora nefesê ya navokî 1 û PGC1 de? tevlihev e û di bin şert û mercên stresê de herî diyar dibe.
Ji bilî vegotina genên ku proteînên mîtokondrial kod dikin, biyojeneya mîtokondrî bi senteza nukleotîdan hewce dike. Çalakkirina genetîkî ya Nrf2 bi rêkûpêkkirina rêça pentose fosfatê û metabolîzma folat û glutamine, nemaze di şaneyên ku bi lez zêde dibin, biosenteza purînê zêde dike (Wêne. 2) [24]. Analîza transkrîptoma Drosophila ya mutant ku ji bo mîtokondrîya serine/treoninîn proteîna kînaza PTEN-ya pîtatîf 1 (PINK1) kêm e, destnîşan kir ku xerabûna mîtokondrial rê li ber verastkirina transkrîpsîyonê ya genan vedike ku bandorê li metabolîzma nucleotîdê dike ku biyolojîk zêde dike [96] mekanîzmayek ji bo parastina li dijî encamên neurotoksîkî yên kêmbûna PINK1 nîşan dide. Nrf2 vegotina fosforîbosil pyrofosfat amidotransferase (PPAT) birêkûpêk dike, ku ketina nav rêça biosyntetîk a nukleotîda de novo purine, û mîtokondrîal methylenetetrahydrofolate dehydrogenase 2 (MTHFD2) katalîz dike (Hêl. 2). Ya paşîn enzîmek bifunctional e bi çalakiyên dehydrogenase û cyclohydrolase ku di peydakirina hem glycine û hem jî formatê de wekî çavkaniyên yekîneyên yek-karbonê ji bo biosenteza purine di şaneyên ku bi lez mezin dibin de girîng e [97]. Ji ber vê yekê îhtîmal e ku aktîvkirina Nrf2 parastî be û dibe ku di kêmasiya PINK1 de xerabûna mîtokondrîal berevajî bike. Bi rastî, aktîvkirina dermankolojîk a Nrf2 ji hêla sulforaphane, an triterpenoid RTA-408, ??m vedigere û hucreyên kêmasiya PINK1 li dijî jehrîbûna dopamînê diparêze [98]. Her çend mekanîzmayên bingehîn tevlihev xuya dikin, bi hev re, ev vedîtin destnîşan dikin ku çalakiya Nrf2 dibe ku bandorê li biyojeneza mitochondrial bike bi bandorkirina asta îfadeya faktorên veguheztinê yên krîtîk û hevaktîfatoran, û her weha bi zêdekirina biosenteza nukleotidê.
Nrf2 û Yekitiya Mînochondrial
Her çend belgeyên yekser hebin hema hema hema hema hema hema hema hema hema hema hema hema hema hema hema hema hema hema hema hema hema hema hema hema hema hema hema hema hema hema hema hema hema hema hema hema hema hebe, heya ku Nrf2 ji bo yekseriya mitochondrial, bi taybetî di mercên oksîterê girîng e. Mitochondria ji ji mizgeftê û muxalîfên giyayê ku biryara yekgirtî ya Nrf2 çalakvanê sulforaphane vekirî ye ku li ser veguherîna pêvajoyê ya mêvotondrialî ya mpotîkelê (mPTP) ji hêla tert-butylhydroperoxide [99], [100] ve hat çêkirin. MPTP, ku komputerê dide ku membrokê mit mitondondrial ê ku bi gelên bi 1500 Da re veşartî bûne, di vê yekê de hate nas kirin ku ji alîgirên F0F1-ATP ve ava kirin [101]. Bi berxwedana sulforaphane-hevpeymaniya mPTP ve bi parastina antioxidant zêdebûna parastin, û asta astengiya GSH, glutathione peroxidase 1, malic enzyme 3, û thioredoxin 2 li hemû heyenên mitochondrial fractions ji alîyê heywanên tedawî yên sulforaphane ve [100].
Zirara proteîna mîtokondrî û xerabûna di nefesê de ku ji hêla hilbera peroksîdasyona lîpîdê ya elektrofîlî 4-hîdroksî-2-nonenal ve hatî çêkirin di mîtokondrîya ku ji kortika mêjî ya mêşên ku bi sulforaphane ve hatî veqetandin ve têne qels kirin [102]. Di hucreyên epîteliya gurçikê yên mişkî de û di gurçikê de, sulforaphane li hember jehrîbûna ji ber cisplatin û gentamicin û windakirina ??m[103], [104] parastin e. Parastina li hember panelek oksîdantan (superoksît, hîdrojen peroksîtê, peroksînîtrite) û elektrofîlan (4-hîdroksî-2-nonenal û acrolein) û zêdebûna parastina antîoksîdan a mîtokondrî jî bi dermankirina hucreyên masûlkeyên nerm ên aorta mişkî bi sulforaphane ve hatine dîtin [105. ]. Di modelek birîna gurçikê ya akût a berevajî de, pêşdibistanên îskemîkî yên lingan di van demên dawî de hate destnîşan kirin ku xwedan bandorên parastinê ye, di nav de astengkirina vekirina mPTP û werimîna mîtokondrial, bi aktîvkirina Nrf2 di encama astengkirina GSK3 de? [106].
Mîtofajî, pêvajoya ku mîtokondrîyên bêfunctional bi bijartî ji hêla otofagozoman ve têne dorpêç kirin û ji lîzozoman re têne şandin ku ji hêla şaneyê ve werin hilweşandin û vezîvirandin, ji bo homeostaza mîtokondrîal girîng e [107], [108]. Digel ku ti têkiliyek sedemî di navbera Nrf2 û mitofagiyê de nehatiye saz kirin, delîl hene ku dibe ku faktora veguheztinê di kontrolkirina kalîteya mitochondrial de bi lîstina di mîtofagiyê de girîng be. Ev dibe ku bi taybetî di bin şert û mercên stresa oksîdative de diyar bibe. Bi vî rengî, di modelek sepsîsê de, zêdebûna astên nîşankera autophagosome MAP1 zincîra sivik 3-II (LC3-II) û proteîna barkêşê p62 di 24 demjimêran de piştî enfeksiyonê di Nrf2-KO de li gorî mişkên WT têne tepisandin [109] . Di van demên dawîn de înduktorek mîtofagiyê ya molekulek piçûk (bi navbeynkariya mitofagiya p62, PMI) hate kifş kirin; ev pêkhateya 1,4-diphenyl-1,2,3-triazole bi eslê xwe wekî çalakkerek Nrf2 hatî çêkirin ku têkiliya faktora veguheztinê bi Keap1 re têk dide [110]. Mîna şaneyên ku tê de Nrf2 ji hêla genetîkî ve hatî rêkûpêk kirin (Keap1-KD an Keap1-KO), şaneyên ku ji PMI re têne xuyang kirin ??m bilindtir in. Ya girîng, zêdebûna herêmîbûna LC3 ya mitochondrial ku piştî dermankirina PMI ya hucreyên WT tê dîtin, di hucreyên Nrf2-KO de çênabe, ku tevlêbûna Nrf2 pêşniyar dike.
Di dawiyê de, analîzasyona ultrastructuralîzasyona liverên liverê diyar kir ku hebûna mestochondrayê bi bi krista kêm kir û membranên di hepatocytes ya Nrf2-KO de, lê ne, Wanê, ku bi hefteyên 24 ve hate xwarinê xwarinê bilind bûye; Bi taybetî, ev gewra delîlên zelal û tevlîheviya oksîdatîk nîşan dide [68]. Ew dikare encam dike ku Nrf2 di çarçoveyek mercên oksîdatîk û tedawî de di binhevkirina mitochondrial de roleke girîng e.
Sulforaphane û Bandorên Wê Li Ser Penceşêr, Mirin, Pîrbûn, Mêjî û Reftarî, Nexweşiya Dil & Zêde
Isothiocyanates hin hinek komên plankirî yên ku hûn di xwarinê de ne. Di vê vîdyoyê de ji bo wan ku ew her dem hatiye çêkirin pir girîng e. Hûrgelan kêmtir? Ji mijara yek ji xalên jêrîn bitikîne bi mijara xwe ya bijartinê vekin. Tiştek temamî bin.
Sernavên sereke
00: 01: 14 - Cancer û mirinê
00: 19: 04 - Aging
00: 26: 30 - Brain and behavior
00: 38: 06 - Reqameya dawî
00: 40: 27 - Dose
Full timeline:
00: 00: 34 - Destpêkkirina sulforaphane, fikirîn sereke ya vîdyoyê.
00: 01: 14 - Gelek mirin û kêmkirina xwarinê hemî mirinê.
00: 02: 12 - rîska kanserê Prostate.
00: 02: 23 - xetereya kansera kanserê.
00: 02: 34 - kansera lûksanê di kişandina xeteran de.
00: 02: 48 - Riskkirina kansera pêsîrê.
00: 03: 13 - Hypothetical: Heke ku hûn berê kanser heye? (interventional)
00: 03: 35 - mekanîzmayên mûzîzalîzmê daneyên danûstendinê yên kanser û mirinê.
00: 04: 38 - Sulforaphane û kanserê.
00: 05: 32 - belgeyên heywanan nîşan bide bandorkirina broccoli nîşan dide ku li ser rîskên tîrêjê di çilê de.
00: 06: 06 - Bandorkirina dermankirinê ya sulforaphane di nexweşxaneyên prostatê de.
00: 07: 09 - Biofilîzasyonê ya metotolyona di tissueya rastîn de.
00: 08: 32 - Pêşangeha kanserên stenbolê stem cells.
00: 08: 53 - Dersa Dîroka: Brassicas wekî xaniyên tendurustî jî di Romayê kevnar de ava kirin.
00: 24: 14 - Xwendekarên Mouse-ê pêşniyar dike ku sulforaphane dibe ku karibe parastinê ya di temenê temenê de çêtirîn.
00: 25: 18 - Sulforaphane pêşveçûna porê çêtirîn di modela mîkrobê de balê. Wêne li 00: 26: 10.
00: 26: 30 - Destpêkkirina mêjû û tevgerên sektorê.
00: 27: 18 - Bandora tesîrê ya broccoli derkeve ser autism.
00: 27: 48 - Bandora glucoraphanîn li schizophrenia.
00: 28: 17 - Destpêkiya dîskêşiyê (mekanîzmayên mektezîzmê û xwendevanan).
00: 31: 21 - Xwendina Mûxoyê bi karanîna xnumxên cuda cuda cuda yên tundûtûjî-stêrdekirin nîşan dide ku sulforaphane bi awayekî fluxoxetine (prozac).
00: 32: 00 - Lêkolînek rasterast ya glucoraphanîn lêkolînek di heman demê de di heman demê de di heman demê de deverek bi bandorkirina astengiya ji modela stratejiya sosyal ve.
00: 33: 01 - Destpêkirina beşa neurodegenerasyon.
00: 33: 30 - Nexweşiya Sulforaphane û Alzheimer.
00: 33: 44 - Nexweşiya Sulforaphane û Parkinson.
00: 33: 51 - Sulforaphane û nexweşiya Hungtington.
00: 35: 01 - Sulforaphane lezgîn piştî ku TBI ji bo bîranîn (xwendina mûsa) çêtir dike.
00: 35: 55 - Sulforaphane û plastîkaya neuronal.
00: 36: 32 - Sulforaphane hînbûna modela şertê duyemîn diabetesê de çêtir dike.
00: 37: 19 - Sulforaphane û duchenne dystrophy.
00: 37: 44 - Myostatin li pêşangehên hucreyên satelê (li vitro).
00: 38: 06 - Daxistina nerazîbûnê û kanserê, zirarê DNA, zexta oksîtatiyê û tehlîzanê, benzene xemgîniyê, nexweşiya cardiovascular, cureyê diyabetê, bandorê li ser mêjî (bandor, xweserîzmê, schizophrenia, neurodegeneration), riya NRF2.
00: 40: 27 - Fikir li ser doktorên broccoli an sulforaphane.
00: 41: 01 - Li ser malbata dorpêçkirin.
00: 43: 14 - Li ser karkirina xwarinê û çalakiya sulforaphane.
00: 43: 45 - Bacteriya guhertinê ya sulforaphane ji glucoraphanin.
00: 44: 24 - karê karên çêtir çêtir dibe gava ku bi tevgera myrosinase ve ji aliyên veberhênanê re hev.
00: 44: 56 - teknolojiyên xwarinê û sebên zêrîn ên xerîb.
00: 46: 06 - Asotiocyanates wek goitrogens.
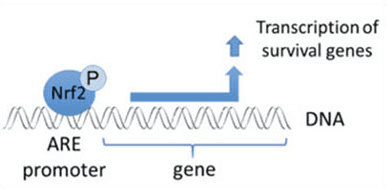
Nrf2 faktoriyek veguherîn e ku sîstema parastina parastina antioxidant a cellorê mirovê mirov a girîng girîng dike. Enerjiya antioxidant elementek an jî, mekanîzmayek rêbazek zagonî ye. Gelek lêkolînên lêkolînan nîşan didin ku NNF2, an jî NF-E2-related factor 2, gelek cûreyên jîndar yên li ser gelek celebên hucreyan veguherîne. Nrf2 jî ji bo parastina hûrgelan û antî-carcinogenicity, ku nîşan dide ku Nrf2 dikare di rêveberiya nexweşî ya neurodegenerative û kanserên ku ji hêla zordariya oksîterî ve têne çêkirin, roleke girîng pêwîste dîtin. Dr. Alex Jimenez DC, CCST Insight
Şîrovekirinên dawî
Her çend gelek pirsan hîn hîn vekirî ye, şahidên ceribandî yên berbiçav eşkere dike ku Nrf2 ê ku di parastina mêvotrojî ya mitochondrial û yekseriya stratejîk de girîng e. Ev rola taybetî di rewşên oksîdative, elektrofîlîk û tedawiya tevgerê de dibe ku meriv bersivkirina Nrf2-bersivên cytoprotîk ên navendî yên navendî û tendurustî bandora tevlihev û organîzmê bandor dike. Roja Nrf2 di fonksiyonê de mitochondrial veguhestineke din a firehrotîkî yên pîvanê vekirî ye ku ji hêla faktora veguherîna vê veguherînê ve tête anîn. Gava ku rewşên pîvanolojîk gelemperî tehlîzasyonê, tehlîmasyon û daketina mitokondrial heye, wekî beşên bingehîn yên pisogenesis, çalakiya dermatolojîk ya Nrf2 ji bo pêşniyarî û dermankirinê ji bo sozê dide. Agahdariya berfirehtir ya mekanîzmayên rastîn ên ku Nrf2 bandor dike bandor dike ku fonksiyonê mitokondrial bandorek ji bo tehlên klînîk ên paşerojê girîng e û dibe ku ji bo bandorên dermankirinê dermankirina dermanên biyomarker pêşkêş dikin.
Armanca gotara li jor nîqaşkirina-û hem jî nîşana-rola derketina Nrf2 di fonksiyona mîtokondrî de bû. Nrf2, an faktoriya nukleer erythroid 2-related factor, rêgezek pêşkeftî ya berxwedana hucreyî ya ji oksîdantan re ye ku dikare beşdarî stresa oksîdasyonê bibe, bandorê li fonksiyona hucreyî bike û bibe sedema pêşveçûna jehrê, nexweşiya kronîk, û tewra kanserê. Dema ku hilberîna oksîdan di laşê mirovan de dikare ji bo armancên cûrbecûr bixebite, di nav de dabeşkirina hucreyê, iltîhaba, fonksiyona berevaniyê, otofajî, û bersiva stresê jî, pêdivî ye ku meriv hilberîna wan zêde were kontrol kirin da ku pêşî li pirsgirêkên tenduristiyê bigire. Qada agahdariya me bi pirsgirêkên tenduristiya chiropractîk û spinal ve sînorkirî ye. Ji bo nîqaşkirina mijarê, ji kerema xwe ji Dr. Jimenez bipirsin an bi me re têkilî daynin915-850-0900.
Êşa piştê. Ew li seranserê cîhanê li seranserê cîhanê sedemên seqetî û rojên bêriya xebatê ye. Painşa piştê sedema duyemîn a herî hevpar a serdanên ofîsa doktor dide, tenê ji hêla enfeksiyonên nefesê yên jorîn zêdetir e. Nêzîkî ji sedî 80yê nifûsê di jiyana xwe de herî kêm carekê êşa piştê dikişîne. Spine avahiyek tevlihev e ku ji hestî, lebat, lebat û masûlke, di nav lebatên nerm ên din de pêk tê. Ji ber vê yekê, birîndarbûn û / an mercên giran, wek dîsên herniated, di dawiyê de dikare bibe sedema nîşanên êşa piştê. Birîndariyên werzîşê an birînên qezaya otomobîlan bi gelemperî sedemên herî êşên piştê ne, lêbelê, carinan tevgerên herî hêsan dikarin encamên êşdar çêbikin. Bi kêfxweşî, vebijarkên dermankirinê yên alternatîf, wekî lênihêrîna chiropractic, dikare bi karanîna verastkirinên stûn û manîpulasyonên destan re êşa piştê sivik bike, di dawiyê de başkirina êş.
Nrf2 piştevanîya çalakiya komên antioxidant û enzymes û genes ku tesîrê mirovê ji bandorên tendurustiyê yên bi zêdebûna asta oxaxative stressê biparêze, wekî nexweşiya Alzheimer biparêzin. Cûreyek cihekî sirûştî ji bo çalakiya Nrf2 veguherandin, ku dikare alîkariya nîşanên nerdejenerative neurodegenerative bikêr bike. Armanca jêrîn li jêr e ku behsa rola sereke ya Nrf2 dibe sedema sedemên zîndanê.
Abstract
Iltîhaba taybetmendiya herî gelemperî ya gelek nexweşî û tevliheviyên kronîk e, dema ku di kanserojenê de rolek krîtîk dilîze. Gelek lêkolînan destnîşan kirin ku Nrf2 bi organîzekirina berhevkirina hucreyên înflamatuar û birêkûpêkkirina vegotina genê bi hêmana bersivê ya antîoksîdan (ARE) beşdarî pêvajoya dijî-înflamatuar dibe. Keap1 (proteîna têkildar a ECH-ê ya mîna Kelch) / Nrf2 (NF-E2-girêdayî faktora 45-ê ya p2) / ARE bi giranî vegotina gena dijî-înflamatuar birêkûpêk dike û pêşveçûna iltîhabayê asteng dike. Ji ber vê yekê, nasîna fîtokîmyasalên dijî-înflamatuar ên nû-girêdayî Nrf2 di vedîtina derman de bûye xalek bingehîn. Di vê vekolînê de, em endamên rêça sînyala Keap1/Nrf2/ARE û genên wê yên jêrîn, bandorên vê rêyê li ser modelên heywanan ên nexweşiyên înflamatuar, û bi riya NF-?B re nîqaş dikin. Digel vê yekê em li ser rêziknameya înflamasoma NLRP3 ji hêla Nrf2 ve jî nîqaş dikin. Ji xeynî vê, em senaryoya heyî ya pêşkeftina fîtokîmîkalên dijî-înflamatuar û yên din ên ku navbeynkariya riya nîşana Nrf2 / ARE dikin kurt dikin.
Infeksiyonê pêvajoya tevlihev e ku ev dibe ku dema hestiyên vexwendiyên zirarên zirarê yên ku bi pîvanogen, zirarê, an dijberî têne birîndar kirin. Hucreyên immune, xweyên xwînê, û navbeynkarên molekular in vê bersiva parastinê [1] hene. Infeksiyonê jî fenomenalek pisîkolojî ye ku bi dewletên cûreyek nexweşiyên ku bi taybetî ji hêla fîzîkî, kîmyewî, biyolojîk û û psîkolojîk ve girêdayî ye. Armanca enfeksiyonê e ku sedema sedemên zirarê, hucre û / an jî hucreyên nerrotîk û hestî yên hûrgelan bisekinin û qedexekirin, û tamandina tissue destpê bike. Du awayên cuda yên inflammasyon cuda ye: zû û zengîn. Sûdema acontasyona xweseriya xwe ya sînor e û sûd e ku meriv mêvandor e, lê dibe sedema sedsala kevneşopî dirêj e ku taybetmendiya gelek nexweşiyên kevneşopî û tehlîlan e. Hêzên dakêşînan ji gelek hucreyên parastinê yên mononîk ên wekî monocytes, macrophages, lîmphocytes û hucreyên plasma, û herweha hilberîna cytokinesê, dibe sedema sedemên zîndanê. Ew tête naskirin ku sermayona zronicî di carcinogenesis [2] rolek girîng e. Bi gelemperî, her du herdu û rêyên şermezar ên germî yên di pêvajoyê de tevlihevkirina tevgerê normal.
Di pêvajoya înflamatuar a patholojîk de, hucreyên mast, monosît, makrofage, lîmfosît û hucreyên din ên parastinê pêşî têne çalak kirin. Dûv re hucre li cîhê zirarê têne peyda kirin, di encamê de celebên oksîjenê yên reaktîf (ROS) çêdibin ku zirarê didin makromolekulan tevî DNA. Di heman demê de, van hucreyên înflamatuar jî mîqdarên mezin navbeynkarên înflamatuar ên wekî cytokines, chemokines, û prostaglandins hilberînin. Van navbeynkaran bêtir makrofajan li deverên herêmî yên iltîhabayê vedigirin û rasterast kaskên veguheztina sînyala pirjimar û faktorên transkripsiyonê yên ku bi iltîhaba re têkildar in çalak dikin. NF-?B (faktora nukleer kappa B), MAPK (proteîna kînaza aktîfkirî ya mîtojen), û JAK (janus kinase) -STAT (veguhezerên sînyalê û aktîvkerên transkrîpsiyonê) rêyên îşaretkirinê di pêşkeftina riya klasîk a iltîhabayê de beşdar in. [3], [4], [5]. Lêkolînên berê diyar kirin ku faktora transkripsiyonê Nrf2 (Faktora 2-ê bi p45-ê ve girêdayî NF-E2) vegotina enzîmên detoksîfker ên qonaxa II-ê di nav de NADPH, NAD(P)H quinone oxidoreductase 1, glutathione peroxidase, ferritin, heme oxygenase-1 ( -1), û genên antîoksîdan ku hucreyan ji birînên cihêreng bi bandorên xwe yên dijî-înflamatuar diparêzin, bi vî rengî bandorê li qursa nexweşiyê dikin [6], [7], [8].
Hişyariya van xuyaniyên girîng, pêşveçûna dermanên dîktatorî yên ji hêla nexweşiyên veguhestinê ve bi rêya rêyên veguhestinê ve di salên dawî de pir zêde balkêş bûn. Di vê nirxandinê de, em lêkolînek li Keap1 (protein re girêdayî Kelê-ê-ECH) / Nrf2 (NF-E2 p45-ê faktor 2) / ARE (reaksiyonek antioxidant reklama) di inflammasyonê de nîşan bide.
Avakirin û Biryara Nrf2
Keap1-Dependent Nrf2 Regulation
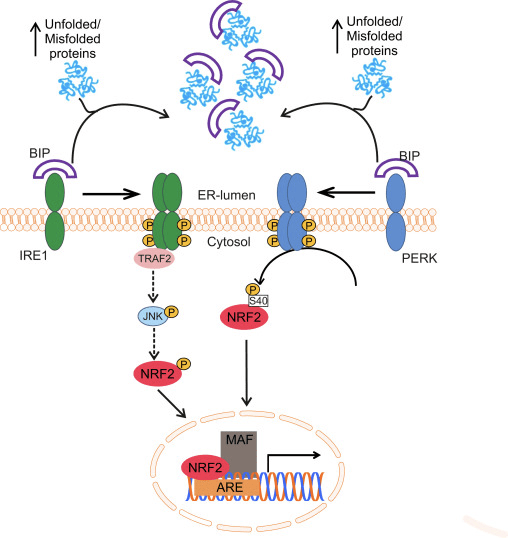
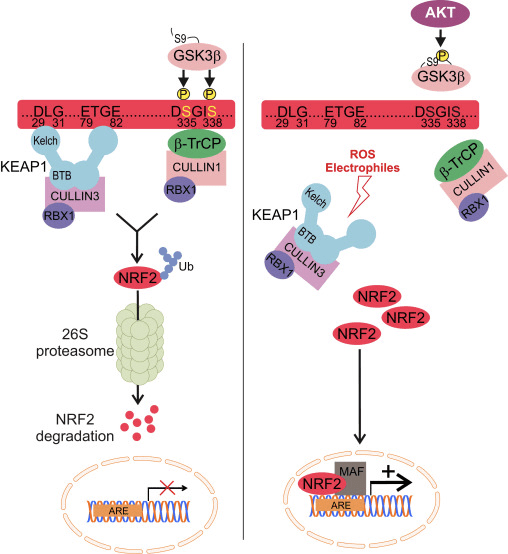
Nrf2 ji binmalbata Cap �n� Collar (CNC) ye û ji heft domên fonksiyonel, Neh (Homolojiya Nrf2-ECH) 1 heya Neh7 [9], [10] pêk tê. Neh1 domanek CNC-bZIP e ku dihêle Nrf2 bi proteînên piçûk ên fîbrosarkoma musculoaponeurotic (Maf), DNA, û hevkarên din ên veguheztinê re heterodimerîze bibe û hem jî bi enzîma UbcM2-ê ku bi ubiquitin-conjugating re kompleksek nukleerî çêbike [11], [12]. Neh2 du motîfên girîng hene ku wekî DLG û ETGE têne zanîn, ku ji bo danûstendina di navbera Nrf2 û regulatorê wê yê neyînî Keap1 de bingehîn in [13], [14].
Keap1 ji bo şertên bingehîn ya CIL-E3 ubiquitin, ji bo şertên veguherînal ên Nrf2 di binpêkirin û proteasomal de di bin şertên normal de [15], [16], [17] de berbi adapter. Destûra KELCH'ê ya Keap1-ê bi hevalên DLG û ETGE yên Nîtrojna Nrf2-Neh2 di cytosolê de, ku ETGE bi têkiliyek bilindtir dike û DLG wekî xêlek [18] dike. Di bin tedawiya oxaxative an jî li ser veguhastina çalakvanên Nrf2, Nrf2 ji ber ku ji guherîna thiol ya Keş1 cysteine veguhestin ji berhevkirina Nrf1 veguhestin û proteasomal hilweşandina 2 [19]. Piştre Nrf2 di nav nucleus de wergera wergerandin, bi protein Mafên Mafê piçûk, û veguherîvanên KES-ê li Battery-genes (Fig. 1A). Carboxy-terminal of Neh3 bi rêveberiya veguherîkasyonê bi rêvegirtina bi co-activator wekî CHD6 (chromo-ATPase / helicase DNA-binding protein) tê naskirin. [20]. Neh4 û Neh5 jî wekî domên veguherînkirina tevgerê, lê bi hev re wekî hevpeymanek veguherînek din ên wekî CBP (camp-response-element-protein-protein-binding protein-binding) tê naskirin. [21]. Ji bilî, Neh4 û Neh5 bi RAC3 / AIB1 / SRC-3-ê, pêşveçûnê ya ku Nrf2-ê vekirî tête axaftin de zehfek zehfî ye. [22]. Neh5 xwedî nîşanek nukleerî ya nukleer a nukleer a nukleer e ku ji bo qaîdeyên û herêmî ya niştimanî ya Nrf2 pir girîng e [23].
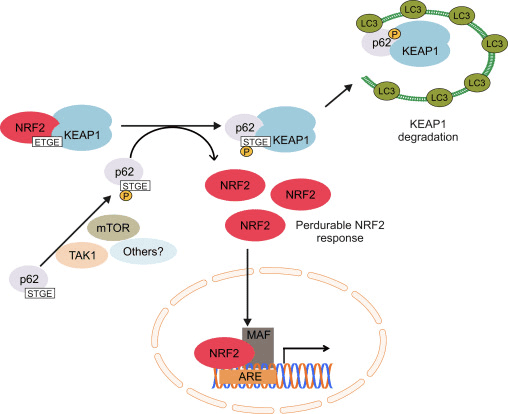
1 Keap1-rexne û girêdayî nexşeya Nrf2. (A) Di bin şert û mercên bingehîn de, Nrf2 bi Keap1 ve ji hêla du motîfên xwe (ETGE û DLG) ve tê veqetandin ku dibe sedema ubiquitination-ya navbeynkar a CUL3 û dûv re hilweşandina proteazomê. Di bin stresa oksîdasyonê de, Nrf2 ji Keap1 vediqete, vediguhezîne navokê û pîlê ARE-gene çalak dike. (B) GSK3 Nrf2 fosforîlate dike û ev yek naskirina Nrf2 ji hêla ?-TrCP ve ji bo ubiquitination bi navbeynkariya CUL1 û dûv re hilweşandina proteazomê hêsan dike. (C) p62 bi Keap1 ve tê veqetandin, ku dibe sedema hilweşîna wê ya otofajîk, azadkirina Nrf2, û nîşana Nrf2 zêde dibe.
Keap1-Qanûna serbixwe Nrf2
Delîlên derketine holê mekanîzmayek nû ya rêziknameya Nrf2 ku ji Keap1 serbixwe ye eşkere kir. Domana Neh6 ya bi serine-dewlemend a Nrf2 di vê rêziknameyê de bi du motîfên xwe (DSGIS û DSAPGS) ve girêdayî bi proteîna ?-transducin-a dubare (?-TrCP) [24] ve, rolek girîng dilîze. ?-TrCP receptorek substratê ye ji bo kompleksa ubiquitin ligase Skp1 Cul1 Rbx1/Roc1 ku Nrf2 ji bo ubiquitination û hilweşandina proteazomê hedef digire. Glycogen synthase kinase-3 proteînek girîng e ku di aramkirin û rêziknameya Nrf1-a serbixwe ya Keap2-ê de têkildar e; ew Nrf2 di qada Neh6 de fosforîlate dike da ku naskirina Nrf2 ji hêla ?-TrCP ve û paşê hilweşîna proteînê [25] hêsantir bike (Wêne. 1B).
Zebûrên din Nrf2
Di heman demê de delîlek din jî diyariya p62-girêdayî çalakiya Nrf2-ê ku di p62-sequesters-deap1-ê dezgehên autofagîk de dibe ku ji dawiyê veguhestinê ya Nrf2 û veguherîna nenasên Nrf2-dependent [26], [27], [ 28], [29] (Nig 1C).
Delîlên berhevkirî destnîşan dikin ku çend miRNA di rêziknameya çalakiya Nrf2 de rolek girîng dileyzin [30]. Sangokoya et al. [31] destnîşan kir ku miR-144 rasterast çalakiya Nrf2 di rêza hucreya lymphoblast K562 de, şaneyên pêşîn ên erythroid mirovan, û retîkulocîtên nexweşiya hucreya dasiyê de birêkûpêk dike. Lêkolînek din a balkêş di şaneyên epîteliya pêsîra mirovî de destnîşan kir ku miR-28 Nrf2 bi mekanîzmayek Keap1-serbixwe [32] asteng dike. Bi vî rengî, miRNAyên wekî miR-153, miR-27a, miR-142-5p, û miR144 di xeta hucreya SH-SY2Y ya neuronal de îfadeya Nrf5 kêm dikin [33]. Singh et al. [34] destnîşan kir ku îfadeya ektopîkî ya miR-93 di modela 2?-estradiol (E17)-ê de ya kanserojeneya mêş de, vegotina genên birêkûpêk ên Nrf2 kêm dike.
Vedîtinek vê dawîyê ji laboratûara me frensek endojen a Nrf2 ku wekî retînoîk X receptor alpha (RXR?) tê zanîn nas kir. RXR? receptorek nukleerî ye, bi domana Neh7 ya Nrf2 (bermayiyên amîno-asîdê 209-316) re bi navgîniya xweya girêdana DNA (DBD) re têkildar dibe, û bi taybetî çalakiya Nrf2 di navokê de asteng dike. Wekî din, receptorên navokî yên din ên wekî receptor-?, ER?, receptor-?-girêdayî estrojenê, û receptorên glucocorticoid-ê yên aktîfkirî yên peroxisome jî hatine ragihandin ku wekî astengkerên endojen ên çalakiya Nrf2 [9], [10].
Roja Nrf2 / HO-1 Axis-Inflammatory
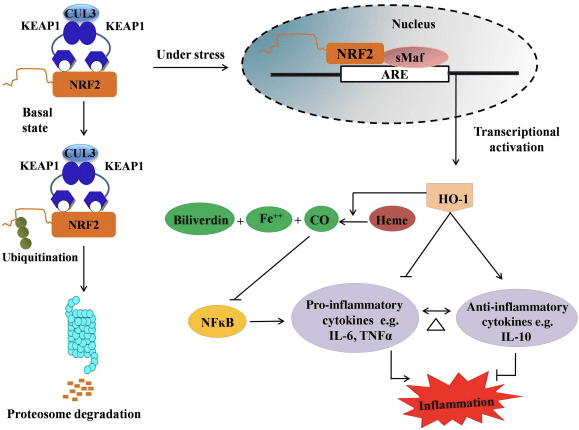
HO-1 di enzyme de hedefê ye û hebûna enzyme-rêjeyê ye ku ew di binavkirina mîkoxoxê (CO) û zêrînek karbonê û bilîbîlîn a bilîbînîn. Hilweşandina enzymatîk ya heme-yê-heme û herweha hilberîna ant-amûranî, wekî CO û bilirubin di warê bandorên parastina HO-1 (Fig. 2) de roleke sereke lîstin.
2 Vebijêrk ya Çavkaniya Nrf2 / HO-1. Di bin şert û mercên bingehîn de, Nrf2 bi represorê xwe Keap1 ve girêdide ku dibe sedema ubiquitination û dûv re hilweşandina proteazomê. Di dema stresa oksîdasyonê de, Nrf2-ya belaş vediguhezîne navokê, li wir bi endamên malbata piçûk a Maf re dimerize û bi genên ARE yên wekî HO-1 ve girêdide. HO-1-ya bilindkirî heme di nav CO, bilirubin û hesinê belaş de katalîze dike. CO wekî astengkerê riya NF-?B tevdigere ku dibe sedema kêmbûna vegotina sîtokînên pro-înflamatuar, di heman demê de bilirubin jî wekî antîoksîdan tevdigere. Digel vê yekê, HO-1 rasterast cytokinesên proînflamatuar asteng dike û hem jî sîtokînên dijî-înflamatuar çalak dike, bi vî rengî rê li ber hevsengkirina pêvajoya înflamatuar vedike.
Nrf2 bi zêdekirina îfadeya mRNA û proteînê gena HO-1 çêdike û ew yek ji genên birêkûpêkkirî yên Nrf2 ye ku bi berfirehî di gelek lêkolînên in vitro û in vivo de tê bikar anîn. Gelek lêkolînan destnîşan kirin ku HO-1 û metabolîtên wê xwedî bandorên girîng ên dijî-înflamatuar ên ku ji hêla Nrf2 ve têne navgîn kirin hene. Bilindbûna derbirîna HO-1 ya ku ji hêla Nrf2-ya aktîfkirî ve tê navbeynkirin rê li ber astengkirina îşaretkirina NF?B dibe sedema kêmbûna birîna mukozê ya rûvî û xerabûna tevnheviyê di modela veguheztina kezeba kezeba nêr Sprague-Dawley [35]. Rêzkirina derbirîna HO-2-girêdayî Nrf1 dibe ku myoblastên C2C12 yên ku ji mişkê hatine wergirtin ji sîtotoxiciya H2O2 biparêze [36]. HO-2-girêdayî Nrf1 bandorek li ser bersivên înflamatuar ên bi navbeynkariya lîpopolîsakarîd (LPS) li RAW264.7- an makrofajên xaneyên kef ên ji makrofajên peritoneal ên mişkî heye. Çalakiya Nrf2 makrofajên hucreya kefiyê fenotîp kêm dike û pêşî li iltîhaba bêserûber a makrofage digire, yên ku di pêşveçûna atherosclerosis de rolek girîng dilîzin [37]. Eksê Nrf2 / HO-1 bandorê li ser hucreyên mîkroglial ên BV2 yên mişkî yên LPS-ê û hucreyên HT22 yên hîpokampal ên mişkî dike, bi bandor li ser neuroînflamasiyonê. Serûberkirina îfadeya HO-1 bi riya Nrf2 di hucreyên mîkroglial ên BV2 yên mişkî de ku mirina hucreyê ya hucreyên hîpokampal HT22 yên mişkî diparêze [38]. Wekî din, molekulên hîbrîd-bingeha kobalt (HYCOs) ku înduktorek Nrf2 bi serbestberek karbonmonoksîtê (CO) re hevûdu dikin, îfadeya Nrf2/HO-1 zêde dikin, CO azad dikin û çalakiya dijî-înflamatuar di vitro de dikin. HYCO di heman demê de HO-1 tevnvîsê sererast dikin û CO-yê di xwînê de piştî rêveberiya di vivo de radigihînin, piştgiriya karanîna wan a potansiyel li dijî şert û mercên înflamatuar [39]. Rêzkirina Nrf2 / HO-1 bi zêdekirina çalakiya efferocytic ya makrofagên mûşî yên ku bi taurine chloramine têne derman kirin iltîhaba kêm dike [40]. Bi tevahî, modelên ceribandî yên ku li jor hatine ravekirin eşkere kirin ku eksê Nrf2 / HO-1 di fonksiyona dijî-înflamatuar de rolek sereke dilîze, û destnîşan dike ku Nrf2 di nexweşiyên bi iltîhaba-girêdayî de armancek dermanker e.
Herweha, ji hêla hilberên HO-1, wekî CO, bilirubin, zordariyek xistî û zirarê hucreyê wekî xeterê xurt dike. [41], [42]; Ew otommune encephalomyelitis û hepatitis teng dike [43], [44]; û ew li dijî hilberîna iNOS û NO [45], [46], [47] bi şikandina endotoxicê mêr û mirinan biparêze. Ji bilî bilî Bilirubin çalakvan û çalakiya endothelial kêm dibe [48]. Bi balkêş e, bilirubin bi rêya transmigration ya leukocytes bi rêya molecule-1-49 [1] kêm dike. Ev referansên taybetî yên nîşanî nîşanên ne tenê-HO-XNUMX wekî agentek antogamatîk-an jî lê jî jî metabolîtên xwe dike.
Medîators û Enzîmên Infeksiyonê yên Nrf2
Cytokines û Chemokines
Cytokines proteînên bi giraniya molekulî û polîpeptîdên kêm in ku ji hêla cûrbecûr şaneyan ve têne derxistin; ew mezinbûna hucreyê, cûdabûn, û fonksiyona parastinê birêkûpêk dikin, û beşdarî iltîhab û başkirina birînan dibin. Sîtokîn înterleukîn (IL), înterferon, faktora nekroza tîmorê (TNF), faktora stimulasyona kolonî, kemokîn, û faktorên mezinbûnê hene. Hin cytokines wekî navbeynkarên pro-înflamatuar têne hesibandin lê yên din xwedî fonksiyonên dijî-înflamatuar in. Zehfkirina stresa oksîdatîf dibe sedema hilberîna zêde ya sîtokînên ku di hucreyên armanc de dibe sedema stresa oksîdative. Dema ku NF-?B ji hêla stresa oksîdatîf ve tê aktîfkirin çend sîtokînên pro-înflamatuar zêde têne hilberandin. Wekî din, stresa oksîdatîf a pro-înflamatuar dibe sedema çalakkirina bêtir NF-?B û hilberîna zêde ya cytokines. Çalakkirina pergala Nrf2/ARE di têkbirina vê çerxê de rolek girîng dilîze. Kemokîn malbatek sîtokînên piçûk in, ku rola wan a sereke rêberkirina koça şaneyên înflamatuar e. Ew bi giranî wekî kemoattractant ji bo leukocît, monocytes, neutrofîl û hucreyên bandorker ên din dixebitin.
Hat ragihandin ku aktîvkirina Nrf2 rê li ber vesazkirina veguheztina LPS-ê ya cytokines pro-înflamatuar digire, di nav de IL-6 û IL-1? [50]. IL-1? û hilberîna IL-6 jî di Nrf2?/? mişkên bi kolîtîya dextran-sulfatê [51], [52]. Nrf2 hilberîna IL-17-a jêrîn û faktorên din ên înflamatuar Th1 û Th17 asteng dike, û pêvajoya nexweşiyê di modelek ceribandî ya skleroza pirjimar, encephalitis otoîmmune [53] de ditepisîne. Genên antî-oksîdan ên girêdayî Nrf2-HO-1, NQO-1, Gclc, û Gclm TNF-?, IL-6, monocyte chemotractant protein-1 (MCP1), macrofage proteîna înflamatuar-2 (MIP2), û înflamatuar asteng dikin. navbeynkaran. Lê di rewşa mêşên Nrf2-knockout de, bandora dijî-înflamatuar pêk nayê [54]. Neutrofîlên peritoneal ên ji mêşên Nrf2-knockout ên ku bi LPS-ê têne derman kirin, ji şaneyên çolê (WT) [6] xwedan astên cytokines (TNF-? û IL-1) û kemokîn (MCP2 û MIP54) pir girîng in [2]. In vitro, veguheztina gena Nrf1 ji hucreyên masûlkeyên nerm ên aortê yên mirov û keroşkê re, derziya MCP8 [55], [2] tepeser dike, û îfadeya HO-1-girêdayî Nrf1, NF-?B û MCP-56-teşwîqkirî TNF-? veşartina di hucreyên endotelyal ên damarê umbilical mirovan de [2]. Van vedîtinan destnîşan dikin ku, di bersiva stimulasyonên înflamatuar de, bilindkirina nîşana NrfXNUMX hilberîna zêde ya sîtokîn û kemokînên pro-înflamatuar asteng dike û her weha çalakkirina NF-?B sînordar dike.
Molekulên Çepê
Molekulên adhezyonê yên şaneyê (CAM) proteîn in ku bi şaneyan an bi matrixa derveyî xaneyê ve girêdidin. Li ser rûyê hucreyê cih digirin, ew di naskirina hucreyê, aktîvkirina hucreyê, veguheztina nîşanê, belavbûn û cûdabûnê de beşdar dibin. Di nav CAM-an de, ICAM-1 û VCAM-1 endamên girîng ên supermalbata immunoglobulin in. ICAM-1 di nav leukocyte û membranên hucreya endothelial de di nav hûrgelên kêm de heye. Li ser teşwîqkirina cytokine, hûrbûn bi girîngî zêde dibe. ICAM-1 dikare ji hêla IL-1 û TNF ve were çêkirin û ji hêla endotelyuma vaskal, makrofage, û lîmfosîtan ve tête diyar kirin. Ew lîgandek întegrînê ye, receptorek ku li ser leukocît tê dîtin. Dema ku pira ICAM-1-integrîn tê çalak kirin, leukocytes bi şaneyên endothelial ve girêdidin û dûv re diçin nav tevnên subendothelial [57]. VCAM-1 navbeynkariya lîmfosîtan, monocytes, eosinophils, û basophils bi endotelyuma vaskal re dike û beşdarî berhevkirina leukocît dike, ku di dawiyê de dibe sedema zirara tevnvîsê ji ber stresa oksîtasyonê. Nrf2 çalakiya promotor a VCAM-1 asteng dike [58]. Gena jêrîn-Nrf2-rêkûpêkkirî HO-1 dikare bandorê li derbirrîna E-selectin û VCAM-1, molekulên adhesionê yên bi hucreyên endothelial re têkildar bike [59]. Ragihandina pişikê ya çend CAM-ên wekî CD-14, TREM1, SELE, SELP, û VCAM-1 di Nrf2?/? mişk ji Nrf2+/+ mişk [60]. Nrf2 di hucreyên endotelyal ên aortê yên mirovî de îfadeya VCAM-1 ya TNF-?-çêkirî ditepisîne û bi girêdana hucreya monosîtîkî ya U937 a TNF-?-ê veguhezîne [8]. Zêde îfadekirina Nrf2 jî di hucreyên endotelyal ên mîkrovaskuler ên mirovî de vegotina gena VCAM-1 a TNF-?-hilweşînkirî asteng dike [61]. Antîoksîdansa xwezayî ya 3-hîdroksantranilic acid (HA), yek ji metabolîtên l-trîptofanê ku di vivo de li ser riya metabolê ku wekî rêça kynurenine tê zanîn di dema iltîhaba an enfeksiyonê de tête çêkirin, tê dîtin ku îfadeya HO-1 derdixe û Nrf2 di umbilical mirovan de teşwîq dike. hucreyên endotelî yên damarê (HUVEC). Vebijandina HO-2-girêdayî Nrf1 ya ku ji hêla HA ve hatî çêkirin, sekreteriya MCP-1, vegotina VCAM-1 û aktîvkirina NF-kB-yê ku bi birîna damaran û iltîhaba di atherosclerosis de ve girêdayî ye asteng dike [56]. Berhevoka chalkone sentetîk a dij-proliferatîf û dij-înflamatuar 2?,4?,6?-tris (metoksymethoxy) chalcone ICAM-1, sîtokina pro-înflamatuar IL-1?, û TNF-? îfadeya di tevna kolonê de ji mêşên ku bi trinitrobenzene sulfonîk asîda hatine derman kirin [62]. Rêzkirina Nrf2 di şaneyên epîteliya pîgmenta retînal a mirovî de ku bi lîkopenê ve têne derman kirin, îfadeya ICAM-1-a ku TNF-?-hilweşînkirî asteng dike [63]. Hemî van lêkolînan destnîşan dikin ku Nrf2 di pêvajoya înflamatuar de bi rêkûpêkkirina koçberî û ketina hucreyên înflamatuar ber bi tevnek iltîhadî ve rolek sereke dilîze.
Matrix Metalloproteinases (MMPs)
MMP bi berfirehî di matrixê derveyî hucreyê de hene û di pêvajoyên fîzyolojîk û patholojîkî yên wekî pirbûna hucreyê, koçberî, cûdabûn, başkirina birînan, angiogenesis, apoptosis, û metastaza tumor de beşdar in. Hat ragihandin ku eksê Nrf2/HO-1 MMP-9 di makrofageyan de û MMP-7 di şaneyên epîteliya rovî ya mirovan de asteng dike, û ev di dermankirina nexweşiya înflamatuar rovî de bikêr e [62], [64]. Zerara çermê ya ku ji ber tîrêjên UV-yê hatî çêkirin di Nrf2-knockout de ji mêşên WT girantir e û asta MMP-9 bi girîngî bilindtir e, ku destnîşan dike ku Nrf2 îfadeya MMP-9 kêm dike. Ji ber vê yekê, Nrf2 wekî parastinê li dijî tîrêjên UV tê hesibandin [65]. Lêkolînek din jî ragihand ku aktîvkirina transkrîpsîyonê ya birêkûpêk a MMP-9 di ketina hucreya tumor û iltîhaba de bi astengkirina riya nîşana NF-kB ve tê rêve kirin [66]. Di birîna stûnê ya trawmatîk de, riya nîşana NF-kB jî di rêzikkirina astên mRNA ya MMP-9 de cih digire [67]. Ji ber vê yekê, di înflamatuar de rêziknameya MMPs rasterast ji hêla riya Nrf2 ve an jî neyekser bi riya NF-?B-bandora Nrf2 ve tê bandor kirin.
Cyclooxygenase-2 (COX2) û Inducible Nitric Oxide Synthase (INOS)
Rêzikek ceribandinên li ser mişkên Nrf2-knockout rola wê ya girîng di iltîhaba û rêziknameya genên pro-înflamatuar ên wekî COX-2 û iNOS de destnîşan kirine. Ji bo cara yekem, Khor et al. rapor zêdekirina cytokinên pro-înflamatuar ên wekî COX-2 û iNOS di tevnên kolonî yên Nrf2?/? mêş bi mêşên WT Nrf2+/+ re hevber kirin, destnîşan dike ku Nrf2 çalakiya wan ditepisîne [51]. Raporek din li ser dermankirina pêşdibistanê bi sulforaphane, yek ji aktîvatorên naskirî yên Nrf2 ku di sebzeyên xaçparêz de heye, bandora wê ya dijî-înflamatuar a astengkirina derbirrîna TNF-?, IL-1?, COX-2, û iNOS li her du mRNA destnîşan kir. û asta proteînê di makrofajên peritoneal ên seretayî yên ji mişkên Nrf2+/+ bi yên ji Nrf2?/? mişk [68]. Bi heman rengî, hîpokampûsa mêşên Nrf2-knockout ên bi iltîhaba LPS-ê vegirtî jî nîşana bilindtirîn nîşankerên iltîhabayê yên wekî iNOS, IL-6, û TNF-? ji mişkên WT [69]. Di heman demê de, mêşên Nrf2-knockout ji stresa oksîdasyonê ya ku ji hêla 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine ve hatî çêkirin, zêde hestiyar in û her weha asta mRNA û proteîna zêde ya nîşankerên iltîhabayê yên wekî COX-2, iNOS nîşan didin. , IL-6, û TNF-? [70]. Wekî din, kezebên ji Nrf2?/? mêşên ku bi parêzek kêmasiya methionine û choline re têkildar in, xwedan ~ 5 qat zêdetir îfadeya mRNA ya Cox2, û iNOS ji yên ji mêşên WT yên li ser heman parêzê ne, rola dijî-înflamatuar ya Nrf2 [71] pêşniyar dikin. Di van demên dawî de, Kim et al. destnîşan kir ku ethyl pyruvate fîtokîmyayî bandorên xwe yên dij-înflamatuar û antî-oksîdative bi kêmkirina îfadeya iNOS-ê bi navgîniya nîşankirina Nrf2 di hucreyên BV2 de dike. Wan destnîşan kir ku ethyl pyruvate veguherîna navokî ya Nrf2 vedigire, ku di dawiyê de pêwendiya di navbera p65 û p300 de asteng dike, û dibe sedema kêmbûna îfadeya iNOS [72]. Wekî din, analoga karbazolê LCY-2-CHO Nrf2 çalak dike û dibe sedema veguheztina wê ya nukleerî, ku rê li ber tepisandina COX2 û îfadeya iNOS [73] di şaneyên masûlkeya xweş a vaskal aortîk a rat de digire.
Rola Paradoksîkî ya Nrf2 Di Rêzkirina Çalakiya NLRP3 iIflammasome de
Malbata NLR, domana pyrin ku 3 (NLRP3) înflamazomê vedihewîne kompleksek pirproteîn e ku wekî receptorek nasîna pathogen (PRR) tevdigere û cûrbecûr nîşaneyên stresê yên mîkrobî, oksîdative yên wekî nimûneyên molekulî yên bi pathogen (PAMPs), zirarê nas dike. molekulên nimûneyên molekulî yên têkildar (DAMP) û ROS [74]. Enflamazoma NLRP3 ya aktîfkirî navbeynkariya perçebûna caspase-1 û derziya cytokine interleukin-1 pro-înflamatuar dike? (IL-1?) ku di dawiyê de pêvajoya mirina hucreyê ku wekî pyroptosis tê zanîn vedigire ku mêvandaran li dijî cûrbecûr pathogenan diparêze [75]. Lêbelê, aktîvkirina nelirêtî ya înflamazomê bi nexweşiyên proteîn ên çewt ên wekî encefalopatiyên spongiform ên veguhezbar, nexweşiya Alzheimer, nexweşiya Parkinson û her weha şekirê celeb 2 [76], kansera [77], gout, û atherosclerosis [78] ve girêdayî ye.
Çavdêriyek vê dawiyê ya koma Rong Hu ya li ser têkiliya Nrf2 bi rêziknameya neyînî ya înflamazomê re eşkere kir ku, Nrf2 îfadeya NQO1 vedigire ku dibe sedema astengkirina çalakkirina înflamazoma NLRP3, perçebûna kaspase-1 û IL-1? nifşê di makrofage de. Wekî din, çalakkerek Nrf2-ya naskirî, tert-butylhydroquinone (tBHQ) bi negatîf veguheztina NLRP3 bi aktîvkirina ARE-yê bi awayê Nrf2-girêdayî [79] ve rêkûpêk kir. Digel çavdêriya jorîn, heman kom di heman demê de hate eşkere kirin ku, dimethyl fumarate (DMF) bi rêya çalakkirina rêça nîşana Nrf2 ya ku di veguheztina nukleerî ya Nrf2 û astengkirina kombûna înflamasome ya NLRP3 de beşdar dibe, pêşî li kolîta ji ber DSS-ê vedike [80].
Rêzikek ceribandinên ku pêkhateyên xwezayî û sentetîk bikar tînin di heman demê de bandora astengker a Nrf2 li ser çalakkirina înflamazoma NLRP3 jî eşkere kiriye. Mînakî, dermankirina epigallocatechin-3-gallate (EGCG) di mişkên lupus nephritis de destnîşan kir ku aktîvkirina înflamazoma NLRP3 ya gurçikê kêm dike ku ji hêla riya nîşana Nrf2 ve tê navgîn kirin [81]. Di heman demê de, citral (3,7-dimethyl-2,6-octadienal), pêkhateyek çalak a sereke di dermanek giyayî ya Chineseînî Litsea cubeba de, çalakkirina înflamazoma NLRP3 bi riya rêça îşaretkirina antîoksîdan a Nrf2 di Nephritîsa Lupus a Lezkirî û Zehmet (ASLN) de asteng dike. [82]. Bi vî rengî, biochanin li hember birîna kezebê ya ji ber LPS/GalN-ê ve hatî çalak kirin bi çalakkirina riya Nrf2 û astengkirina çalakkirina înflamazoma NLRP3 di mêşên BALB/c yên nêr de diparêze [83]. Digel vê yekê, mangiferin di heman demê de hate destnîşan kirin ku vegotina Nrf2 û HO-1 bi rengek bi doz ve girêdayî ye û LPS/D-GalN-ê-xebatî NLRP3, ASC, caspase-1, IL-1 asteng dike? û TNF-? îfade [84].
Tevî rêziknameya neyînî ya NLRP3 ji hêla Nrf2 ve, ew fonksiyona înflamasome ya NLRP3 û AIM2 jî çalak dike. Haitao Wen û hevalên wî vedîtin ku, Nrf2 ?/? Makrofagên mişkî çalakkirina xelet a NLRP3 û AIM2 Inflammasome lê ne înflamazoma NLRC4 nîşan dane [85]. Balkêş e, ev çavdêrî fonksiyonên nenas ên Nrf2 di çarçoweya nexweşiyên bi iltîhaba têkildar de nîşan dide; ji ber vê yekê pir girîng e ku meriv bêtir lêkolîn bike da ku mekanîzmaya ku tê de Nrf2 fonksiyona înflamasome çalak dike eşkere bike berî ku ew wekî armancek dermankirinê were hesibandin.
Zordariya Pro-Inflammatory Transcription Cytokine by Nrf2
Lêpirsînek pir nû ya ku li ser bingeha immunoprecipitation kromatin (ChIP)-seq û ChIP-qPCR di makrofagên mişkî de encam dide, eşkere kir ku Nrf2 bi herêmên pêşverû yên sîtokînên pro-înflamatuar ên wekî IL-6 û IL-1 ve girêdide? û leşkerkirina RNA Pol II asteng dike. Wekî encamek, RNA Pol II nekare aktîvkirina transkrîpsiyonê ya IL-6 û IL-1 pêvajoyê bike? ku di dawiyê de dibe sedema astengkirina vegotina genê. Yekem car, koma Masayuki Yamamoto mekanîzmaya nû eşkere kir ku bi wê Nrf2 ne tenê genên xwe yên jêrîn bi navgîniya AREs veguhezîne lê di heman demê de çalakkirina transkrîpsîyonî ya genên taybetî bi an bêyî ARE-yê bi astengkirina peydakirina RNA Pol II [50] ditepisîne.
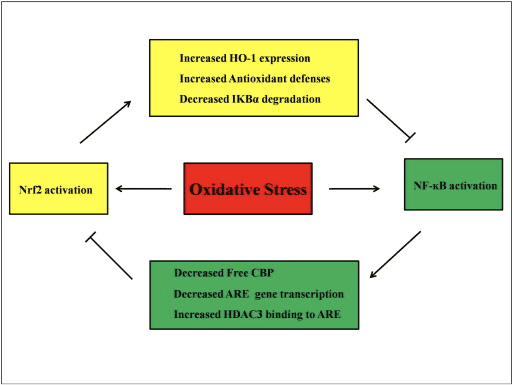
Di Navbera Riyên Nrf2 û NF-?B de Xaçakkirin
NF-?B kompleksek proteîn e ku berpirsiyarê veguheztina DNA-yê ye ku hema hema di hemî celeb şaneyên heywanan de tê dîtin û di pêvajoyên cihêreng ên wekî iltîhaba, apoptosis, bersivdayîna berevaniyê, mezinbûna hucreyê û pêşkeftinê de tê dîtin. P65, proteînek Rel ya ji malbata NF-?B, xwedan domanek transaktîvasyonê ye lê p50 tune ye û ji bo aktîfkirina transkripsiyonê bi proteîna Rel re heterodimerîzasyon hewce dike. Di dema stresa oksîdatîf de, I?B kînaza (IKK) tê çalak kirin û dibe sedema fosforîlasyona I?B, di encamê de berdan û veguheztina navokî ya NF-?B. NF-?B dibe sedema transkripsiyona navbeynkarên pro-înflamatuar ên wekî IL-6, TNF-?, iNOS, IL-1, û adhesiona hundurîn COX-2.
Rêzkirina anormal ya NF-?B bi gewrîta romatoid, astim, nexweşiya înflamatuar a rovî û Helicobacter pylori gastritis ve girêdayî ye [86]. Naha tê hesibandin ku çalakiya NF-kB bi piranî di sê aliyan de bandorê li riya nîşankirina Keapl/Nrf2/ARE dike: yekem, Keap1 IKK-ê xirab dike? bi rêya ubiquitination, bi vî awayî çalakiya NF-?B asteng dike [87]. Ya duyemîn, pêvajoya înflamatuar navbeynkarên înflamatuar ên mîna COX2 ku ji cyclopentenone prostaglandin 15d-PGJ2, elektrofîlek bihêz a ku bi Keap1 re reaksiyonê dike û Nrf2 çalak dike, vedigire, bi vî rengî dest bi veguheztina genê bi astengkirina hevdemî ya çalakiya NF-kB dike [58] Wêne 88 A, B). Ya sêyem, NF-?B dikare bi hev-aktîvkera transkrîpsiyona Nrf3 ya reqabetê ya CBP [2], [89] re bike yek (Hêjî. 90 C, D).
Figure 3 Di navbera rêyên Nrf2 û NF-?B de hevpeyivîn. (A) Keap1 IKK-ê rê dide ubiquitination û hilweşandina proteazomê ya bi navbeynkariya CUL3 ku di dawiyê de dibe sedema astengkirina fosforîlasyona NF-?B û ev mekanîzma di heman demê de wekî girêdana pêşbazî ya Nrf2 û IKK bi Keap1 re dixebite. (B) Stresa oksîdatîf IKK-ya ku NF-?B fosforîlate dike çalak dike, dibe sedema veguheztina wê nav nucleus û aktîvkirina sîtokînên proînflamatuar ên wekî COX-2. Hilbera termînalê ya COX-2 ku wekî 15d-PGJ2 tê zanîn wekî înduktorek Nrf2 tevdigere ku di dawiyê de rê li ber tepisandina stresa oksîdative vedike. (C) Nrf2 bi kofaktora xweya transkrîpsiyona CBP re digel Maf-a piçûk û makîneyên din ên veguheztinê ve girêdide da ku dest bi vegotina genê ya ARE-ê bike. (D) Dema ku NF-?B bi CBP-ê re bi rengek pêşbaziyê ve girêdide, ew girêdana CBP-ê bi Nrf2 re asteng dike, ku dibe sedema astengkirina veguheztina Nrf2.
Tê texmîn kirin ku rêyên nîşana Nrf2 û NF-?B bi hevûdu re têkildar in da ku veguheztin an fonksiyona proteînên hedefa jêrîn kontrol bikin. Di rastdariya vê texmînê de gelek mînak destnîşan dikin ku aktîvkirin û astengkirina rasterast an nerasterast di navbera endamên rêyên Nrf2 û NF-?B de çêdibe (Hêjî. 4). Di bersiva LPS-ê de, têkçûna Nrf2 bi girîngî çalakiya veguheztina NF-?B û veguheztina genê ya girêdayî NF-?B-ê zêde dike, û destnîşan dike ku Nrf2 çalakiya NF-?B asteng dike [60], [91]. Digel vê yekê, zêdekirina vegotina HO-2-ya jêrîn a Nrf1-girêdayî çalakiya NF-?B asteng dike. Dema ku hucreyên penceşêrê yên prostatê bi kurtasî li ber ?-tochopheryl succinate, ku jêderek vîtamîna E, têne xuyang kirin, îfadeya HO-1 zêde dibe. Berhemên dawî yên HO-1 veguheztina navokî ya NF-?B asteng dike [92]. Ev di lêkolînên vivo de pêşniyar dike ku Nrf2 bi rê ve girêdayî rêberê NF-kB bi rê ve dike. LPS çalakiya girêdana DNA ya NF-?B teşwîq dike û asta binyekeya p65 ya NF-?B di ekstraktên nukleerî yên ji pişikên Nrf2?/? de pir zêde ye. ji mişkên WT, rola neyînî ya Nrf2 di çalakkirina NF-?B de pêşniyar dike. Wekî din, Nrf2?/? Fîbroblastên embrîyoya mişkî yên bi LPS û TNF-? çalakkirina NF-?B ya ku ji hêla çalakkirina IKK û I?B-? hilweşandin [60]. Û paqijkirina virusa syncytial respirasyonê bi girîngî kêm dibe dema ku çalakiya girêdana DNA ya NF-?B di Nrf2?/? de zêde dibe. mêş bi mişkên WT re [93]. Nephritiya lupusê ya ku ji hêla Pristane ve hatî çêkirin di Nrf2?/? mêşên ku bi sulforaphane re hevbeş têne derman kirin xwedan zirara gurçikê ya giran û guhertinên patholojîkî ne û her weha li gorî WT-ê îfadeya iNOS û aktîvkirina NF-?B bilindtir e, ku pêşniyar dike ku Nrf2 nefrita lupusê bi astengkirina rêça sînyala NF-?B û paqijkirina ROS [94. ]. Çalakiya NF-?B di heman demê de dema ku hucre bi LPS û TNF-? re bi inducerek Nrf2 re têne derman kirin jî çêdibe. Mînakî, hilberek chalcone ya sentetîk aktîvkirina NF-?B ya ku TNF-?-hilweşiyayî hem rasterast hem nerasterast û hem jî bi qismî bi navgîniya vegotina HO-1 di şaneyên HT-29-a epîteliya rovî ya mirovî de asteng dike [62]. Tepeserkirina veguheztina NF-?B û çalakiya girêdana ADN-ê û her weha tepeserkirina vegotina iNOS di hepatocîtan de dema ku mişkên F344 bi 3H-1,2-dithiole-3-thione (D3T) têne derman kirin [95] têne dîtin. Piştî hev-dermankirina bi sulforaphane û LPS, bilêvkirina LPS-ê ya iNOS, COX-2, û TNF-? di Raw 264.7 de makrofage têne kêmkirin, pêşniyar kirin ku sulforaphane xwedan çalakiya dijî-înflamatuar e bi astengkirina girêdana DNA ya NF-?B [96]. Her çend gelek lêkolînên ezmûnî hatine kirin da ku têkiliya di navbera rêyên Nrf2 û NF-?B de rave bikin, encamên nakokî dimînin. Hemî rêbazên erênî û neyînî di navbera Nrf2 û NF-kB de hat ragihandin. [97]. Bi gelemperî, elektrofîlên kîmopreventive 3H-1,2-dithiole-3-thione, sulforaphane û Triterpenoid CDDO-Me çalakvanên NF-kB û cinsên wê yên xeletî li [2], [98], [99] têkevin. Berevajî, gelek şertên an jî mercên wekî ROS, LPS, zexta hehê, LDL oxoxized, û dûmana cigarê hatine nîşandan ku çalakiya Nrf2 û NF-KB jî zêde dibe. [97]. Wekî din, lêkolînên in vivo diyar kirin ku çalakiya NF-kB di kezebên ji Nrf2?/? veqetandî de kêm dibe kêm dibe. mêş û çalakiya girêdana NF-?B di Nrf2?/? ji mişkên Nrf2+/+ [101]. Lêbelê, hucreyên endotelyal ên aortê yên mirovî yên ku bi vektora adenovîral Nrf2 têne derman kirin genên jêrîn ên NF-?B asteng dikin bêyî ku bandorê li çalakiya NF-?B bike [8].
Figure 4 Loop Rêziknameya Nrf2 û NF-?B. Rêya Nrf2 bi rêgirtina li hilweşîna I?B-? ve çalakkirina NF-?B asteng dike. û zêdekirina îfadeya HO-1 û berevaniya antîoksîdan ku ROS û kîmyewiyên detoksîfker bêbandor dike. Wekî encamek, aktîvkirina NF-?B ya girêdayî ROS-ê tê tepisandin. Bi heman rengî, transkripsiyona NF-?B-navbeynkar bi kêmkirina çalakkirina Nrf2 kêm dike.InVeguheztina genê û proteîna girêdana CREB ya belaş bi pêşbaziya bi Nrf2 re ji bo CBP. Digel vê yekê, NF-?B berhevkirina hîstone deacetylase (HDAC3) li herêma ARE zêde dike û ji ber vê yekê çalakkirina transkrîpsiyona Nrf2 tê asteng kirin.
Çalakiya niştimanî ya Nrf2 di roleke enzîmên û genes de di tevlêbûna oxixantên reaksiyonê de bi hêza antioxidant a hucreyan di bedenê mirovan de tevlihev dike. Dema ku îro lêkolînên gelek lêkolîn hene, îro mekanîzmayên rêbazê di çalakiya Nrf2 de ne temam nayên zanîn. Roja mimkin a rêbazê ya Nrf2 di bin tedawiya tevlîbûnê de hat dîtin. Dr. Alex Jimenez DC, CCST Insight
Role ya Nrf2 di Nexweşên Infeksiyonê de
Di lêkolînên vivo da ku nîşan dide ku Nrf2 di nexweşiyên cûda yên bandor ên bandor ên bandor dike bandorek girîng e; Di nav de gastritis, kolîtîk, germîtis, nehûne, zirarê liver, nexweşî nexweşî, nexweşî û nexweşî. Di van lêkolînan de, Nrf2?/? heywanan ji heywanên WT nîşanên girantir ên iltîhaba û zirara tevnê nîşan dan. Ji ber vê yekê tê guman kirin ku rêgeziya Nrf2 bi bandorkirina tedawiya parastinê heye. Sazkirina intra-tracheal ya elastase ya pancreatê bi astengiya peproşokî, bi taybetî jî emphysema hilberîne. Nîşaneyên Nrf2-ê ji bo emphysema gelemperî ne, û xuyabûna HO-1, PrxI, û gene antiprotease SLPI di nav alveolar macrophages de têne kêm kirin. Nrf2 li ser pergala parastina navberî ya li dijî zirara laşê [102] tête navîn a keyfêkerê serekî ye. Bişkojka Nrf2-êkêşî bi emphysema ku ji bo 6-ê vexwarinê veguhestina tîrêjê ya tutunê nîşan dide, dihêlehejîna bronchoalveolar nîşan dide, zelalkirina rêjeya oksîdative stressên di alveoli de, û apoptosê ya seopotîk a sevalîtal zêde bû, pêşniyar dike ku Nrf2 li hemberî enfixîdantiya zêdebûna tutunê tutunê tutunê dike. genes [102], [103]. Li gel Nrf2 bi astengî, tevliheviya avê ya allergen-mediated û astengkirina paqijkirina ovalbuminê ya avê, hewaya hyper-reaktîvî, hîperplasia ya goblet, û asta Th2 di pergala bronchoalveolar de bronchoalveolar di nav avê de hewaya eosinophilia bilind dike. , mîkrobek hîderkêşanê, û avêtina hîpera hyper û herweha gelek cinsên antioxidant ên ku pêşveçûna Asthma [2] çêbikin. Înfîlîzê ya Carrageenan di nav kûrahiya dilovaniyê de desthilatdar dike, û kombûnên 15d-PGJ2 di hucreyên tevgerê yên Nrf2 de bi mîkrobatên peritoneal ve girêdayî ye. Di dema pêvajoya destpêkê de, 15d-PGJ2 Nrf2 çalak dike û pêvajoya tevlîbûna HO-1 û PrxI bi pêvajoya tevlihevkirinê veguherîne. Lêkolînek jî diyar kir ku COX-2 di pêvajoya destpêkê de ji hêla hilberîna 15d-PGJ2 ve [105]. Rêveberiya devkî ya 1% dextran sulfate sodium ji bo 1 hefte bi kulîteyên têkildar yên bi hîstolojî ve girêdayî ye ku hûrgelên kurt û rûxandina hucreyên germî yên li kolonê. Ji bo parastina integrasyonê li Kolîteyê, Nrf2 dikare bi hêla rêbaziya pro-inflammatory cytokines re dikeve û pêvajoya duyemîn ê enzyme detoxifying [xNUMX] dikir. Di modelek mişkek Nrf2-knockout ya sepsîsa pişikê ya ku ji hêla LPS ve hatî çêkirin de, çalakiya NF-?B bandora sîtokînên înflamatuar ên wekî COX-2, IL-113, IL-6, û TNF? ku ji bo destpêkirin û pêşvebirina iltîhaba bingehîn in [60]. Nrf2 ji hêla faktorên devokrasiyê veguherandina zirarê germê kêm dike. Di van modelan de hejmarê acute, enfeksiyonên zêdebûna enzymes, enfeksiyonên pro-agamatorî, û navbeynkarên bi Nrf2 riya nîşankirina Nerazîbûna WT-ê di WT heywanan de kêm dike. Bi balkêş e, ev di heman demê de di heman demê de di heman demê de di heman demê de ji bo Nîşaneyên NTF2-ê ve hatine ragihandin ku di navnîşanên WT-ê de bi nîşanên nerazîbûnê têne zelal kirin.
Lêkolînê li Nrf2-Dependent Anti-Inflammatory Drugs
Di encam de, em tecrûbeyên ku di rêgeziya êşanînê de Nelf2 veguherînek rêbazek rêbazên lîstikvan dike, ji ber ku nenasên nrf2-dependent ji bo tedawîkirina tedawiya nermaletê girîng e.
Plants çavkaniyên bêhtir çavkaniyên dewlemend ên ku ji hêla veguherîna Nrf2 ve tê veşartin, ku ji bo jinan cytoprotective up-up. Di dawiyê de, gelek lêkolînan pêk hat ku bandorên bandorên antogammatîk yên piranî, piranîya nebatên hilberînê lêkolîn bikin. Ji bo nimûne, curcumin celebek çalakiyek tundir e û di piçûkek piçûk de di nav deynê de ye. isotiocyanates, bi taybetî bi fenylisothiocyanates ji brookoli, gemiyê, û aliyên din hene; û anthocyanins ji ber berazan û gurên têne [124]. Lêkolînan nîşan kir ku ev hemû nûnerên ne tenê antioxidants ne, lê bi bandorên enfeksiyonê bi rêya Nrf2-ê [125], [126] jî xwedî hêzdar heye. Ji ber vê yekê, pêşveçûna çalakvanên ant-inflammatory Nrf2 veguhestin ji birîna çandiniyê vekolînek di lêkolîna bijîşkek pir zêde.
Di salên dawî de, gelek ceribandinên heywanan hatine kirin ku kiryarên van pêkhateyan piştrast bikin. Artesunate bi giranî ji bo malariya giran, malariya mejî, û nexweşiyên otoîmmune romatîkî tê bikar anîn; di birîna pişikê ya septîk de jî bi bandor e. Artesunate îfadeya Nrf2 û HO-1 çalak dike, û ya paşîn ketina cytokines û leukocît pro-înflamatuar di nav tevnê de kêm dike da ku pêşî li iltîhaba bigire [127]. Isovitexin, ku ji qalikên birincê Oryza sativa tê derxistin, tê fikirîn ku xwediyê taybetmendiyên dijî-înflamatuar û antîoksîdan e; ew bi çalakkirina riya Nrf2/HO-1 û astengkirina MAPK û NF-?B [128], li hember birîna pişikê ya akût a ku ji hêla LPS ve hatî çêkirin de rolek parastinê dilîze. Fimasartan, astengkerek receptorê angiotensin II ya nû populer ku li ser pergala renin-angiotensin tevdigere, tansiyona xwînê kêm dike; bikaranîna fimasartan ji bo dermankirina mişkan bi astengiya ureteral a yekalî ya bi emeliyatê ve, stresa oksîdative, iltîhaba, û fibrosisê bi rêgezkirina Nrf2 û riya antîoksîdan û astengkirina RAS û MAPKs kêm dike [129]. Sappanone bi berfirehî li Asyaya Başûr-rojhilatê tê belav kirin, ku li wir wekî dermanek dijî-înfluenza, antî-alerjîk û neuroprotektîf tê bikar anîn; ew Nrf2 çalak dike û NF-?B asteng dike û ji ber vê yekê dibe ku di dermankirina nexweşiyên Nrf2- û/an NF-?B-yên têkildar de sûdmend be [130]. Bixina ku ji tovên Bixin orellana tê derxistin li Meksîka û Amerîkaya Başûr ji bo nexweşiyên infeksiyonî û înflamatuar tê bikaranîn; ew navbeynkarên înflamatuar, rijandina kapîlara alveolar, û zirara oksîtasyonê bi rengek girêdayî Nrf2 kêm dike da ku birîna pişikê ya ku ji hewa veguhezîne sivik bike û morfolojiya pişikê ya normal vegerîne [131]. Pêkhateyên nebatê yên din, wek epigallocatechin gallate, sulforaphane, resveratrol, lycopene, û jêgirtina çaya kesk bi riya riya nîşana Nrf2 re bandorên dermankirinê li ser nexweşiyên înflamatuar hene [132], [133], [134]. Di van demên dawîn de fîtokîmyakî din, erodictyol, ku di fêkiyên citrus de heye, hate ragihandin ku bandorên antî-înflamatuar û antîoksîdan li ser birîna gurçikê ya ji cisplatin-ê û birîna pişikê ya akût a ku ji ber sepsîsê ve hatî çêkirin heye bi rêkûpêkkirina Nrf2, astengkirina NF-?B û astengkirina îfadeya sîtokinan di makrofageyan de [135], [136]. Lêbelê, gelek fîtokîmyayî ji bo pêşîlêgirtin û dermankirina cûrbecûr nexweşiyên mirovî sozek mezin nîşan didin, û hin jî berê ketine qonaxa ceribandinên klînîkî (Table 2).
Ev cilên mektebên ku di rêgezên nerazîbûna Nrof2 de bi awayekî materyalê elektrofîlîk re veguherînin ku ji kapîtalên cysteine yên Keap1 veguherînin, ji bo ku bi rêkûpêk Nrf2-ê ve girêdayî ye.
Sulforaphane û Bandorên Wê Li Ser Penceşêr, Mirin, Pîrbûn, Mêjî û Reftarî, Nexweşiya Dil & Zêde
Isothiocyanates hin hinek komên plankirî yên ku hûn di xwarinê de ne. Di vê vîdyoyê de ji bo wan ku ew her dem hatiye çêkirin pir girîng e. Hûrgelan kêmtir? Ji mijara yek ji xalên jêrîn bitikîne bi mijara xwe ya bijartinê vekin. Tiştek temamî bin.
Sernavên sereke
00: 01: 14 - Cancer û mirinê
00: 19: 04 - Aging
00: 26: 30 - Brain and behavior
00: 38: 06 - Reqameya dawî
00: 40: 27 - Dose
Full timeline:
00: 00: 34 - Destpêkkirina sulforaphane, fikirîn sereke ya vîdyoyê.
00: 01: 14 - Gelek mirin û kêmkirina xwarinê hemî mirinê.
00: 02: 12 - rîska kanserê Prostate.
00: 02: 23 - xetereya kansera kanserê.
00: 02: 34 - kansera lûksanê di kişandina xeteran de.
00: 02: 48 - Riskkirina kansera pêsîrê.
00: 03: 13 - Hypothetical: Heke ku hûn berê kanser heye? (interventional)
00: 03: 35 - mekanîzmayên mûzîzalîzmê daneyên danûstendinê yên kanser û mirinê.
00: 04: 38 - Sulforaphane û kanserê.
00: 05: 32 - belgeyên heywanan nîşan bide bandorkirina broccoli nîşan dide ku li ser rîskên tîrêjê di çilê de.
00: 06: 06 - Bandorkirina dermankirinê ya sulforaphane di nexweşxaneyên prostatê de.
00: 07: 09 - Biofilîzasyonê ya metotolyona di tissueya rastîn de.
00: 08: 32 - Pêşangeha kanserên stenbolê stem cells.
00: 08: 53 - Dersa Dîroka: Brassicas wekî xaniyên tendurustî jî di Romayê kevnar de ava kirin.
00: 24: 14 - Xwendekarên Mouse-ê pêşniyar dike ku sulforaphane dibe ku karibe parastinê ya di temenê temenê de çêtirîn.
00: 25: 18 - Sulforaphane pêşveçûna porê çêtirîn di modela mîkrobê de balê. Wêne li 00: 26: 10.
00: 26: 30 - Destpêkkirina mêjû û tevgerên sektorê.
00: 27: 18 - Bandora tesîrê ya broccoli derkeve ser autism.
00: 27: 48 - Bandora glucoraphanîn li schizophrenia.
00: 28: 17 - Destpêkiya dîskêşiyê (mekanîzmayên mektezîzmê û xwendevanan).
00: 31: 21 - Xwendina Mûxoyê bi karanîna xnumxên cuda cuda cuda yên tundûtûjî-stêrdekirin nîşan dide ku sulforaphane bi awayekî fluxoxetine (prozac).
00: 32: 00 - Lêkolînek rasterast ya glucoraphanîn lêkolînek di heman demê de di heman demê de di heman demê de deverek bi bandorkirina astengiya ji modela stratejiya sosyal ve.
00: 33: 01 - Destpêkirina beşa neurodegenerasyon.
00: 33: 30 - Nexweşiya Sulforaphane û Alzheimer.
00: 33: 44 - Nexweşiya Sulforaphane û Parkinson.
00: 33: 51 - Sulforaphane û nexweşiya Hungtington.
00: 35: 01 - Sulforaphane lezgîn piştî ku TBI ji bo bîranîn (xwendina mûsa) çêtir dike.
00: 35: 55 - Sulforaphane û plastîkaya neuronal.
00: 36: 32 - Sulforaphane hînbûna modela şertê duyemîn diabetesê de çêtir dike.
00: 37: 19 - Sulforaphane û duchenne dystrophy.
00: 37: 44 - Myostatin li pêşangehên hucreyên satelê (li vitro).
00: 38: 06 - Daxistina nerazîbûnê û kanserê, zirarê DNA, zexta oksîtatiyê û tehlîzanê, benzene xemgîniyê, nexweşiya cardiovascular, cureyê diyabetê, bandorê li ser mêjî (bandor, xweserîzmê, schizophrenia, neurodegeneration), riya NRF2.
00: 40: 27 - Fikir li ser doktorên broccoli an sulforaphane.
00: 41: 01 - Li ser malbata dorpêçkirin.
00: 43: 14 - Li ser karkirina xwarinê û çalakiya sulforaphane.
00: 43: 45 - Bacteriya guhertinê ya sulforaphane ji glucoraphanin.
00: 44: 24 - karê karên çêtir çêtir dibe gava ku bi tevgera myrosinase ve ji aliyên veberhênanê re hev.
00: 44: 56 - teknolojiyên xwarinê û sebên zêrîn ên xerîb.
00: 46: 06 - Asotiocyanates wek goitrogens.
encamên
Heya nuha, pir lêkolîn li ser rola rêça nîşana Nrf2 / Keap1 / ARE di iltîhaba de sekinîne. Di nav enzîmên ku ji hêla Nrf2 ve têne rêve kirin de, HO-1 yek ji wan enzîmên bersiva stresê nûner e. HO-1 xwedan taybetmendiyên antî-înflamatuar û antîoksîdan ên berbiçav e. Bi gelemperî, riya nîşana Nrf2 di heman demê de cytokines, faktorên berdana kemokîn, MMPs, û navbeynkarên din ên înflamatuar COX-2 û hilberîna iNOS-ê jî neyînî bi rê ve dibe, ku rasterast an nerasterast bandorê li rêyên NF-kB û MAPK-ê yên têkildar û torên din ên ku iltîhaba kontrol dikin dike. Tê pêşnîyar kirin ku rêyên nîşana Nrf2 û NF-?B bi hevûdu re têkilî daynin da ku veguheztin an fonksiyona proteînên hedefa jêrîn birêkûpêk bikin. Tepeserkirin an neçalakkirina çalakiya transkrîpsiyonê ya bi navbeynkariya NF-?B bi navbeynkariya Nrf2 bi îhtîmalek mezin di qonaxa destpêkê ya iltîhabayê de pêk tê, ji ber ku NF-?B senteza de novo ya komek navbeynkarên pro-înflamatuar bi rê ve dibe. Lêbelê, hîn jî di lêkolînê de hin sînor hene, wek mînak gelo girêdanên di navbera Nrf2 û rêyên nîşankirinê yên din ên wekî JAK/STAT de hene, girîngiya aktîvatorên Nrf2 yên heyî yên ku ji çavkaniyên nebatên xwezayî yên di iltîhabayê de têne peyda kirin, û meriv çawa çalakiya biyolojîkî baştir dike. û armanckirina van pêkhateyan zêde bike. Vana erêkirina ceribandinê ya bêtir hewce dike.
Wekî din, riya nîşana Nrf2 dikare> 600 genan [163] birêkûpêk bike, ji wan > 200 proteînên sîtoprotektîf [164] kod dikin ku di heman demê de bi iltîhaba, kansera, nexweşiyên neurodejenerative, û nexweşiyên din ên sereke re têkildar in [165]. Zêdebûna delîlên ku destnîşan dikin ku, riya nîşana Nrf2 di gelek penceşêrê de bêrêkûpêk e, ku di encamê de battêra genê ya girêdayî Nrf2-ê bi derbirrînek bêserûber derdikeve. Digel vê yekê, iltîhaba di nexweşiyên têkildarî stresa oksîdative bi taybetî di kanserê de rolek sereke dilîze. Serîlêdana çend aktîvatorên Nrf2 ji bo berevajîkirina iltîhabayê dibe ku bibe sedema vegotina nelirêtî ya genên jêrîn ên Nrf2 ku onkogenesis û berxwedana li dijî kemo û/an terapiya radyoyê çêdike. Ji ber vê yekê, çalakkerên pir taybetî yên Nrf2 dikarin werin pêşve xistin da ku bandorên wê yên pleiotropîk kêm bikin. Gelek çalakvanên Nrf2 di nexweşiyên têkildarî stresa oksîdative de çêtirbûnek girîng a fonksiyonên dijî-înflamatuar nîşan dane. Mînaka çêtirîn a aktîvatorê Nrf2 ku ji hêla FDA ve hatî pejirandin û bi berfirehî ji bo dermankirina nexweşiya înflamatuar wekî skleroza Pirjimar (MS) tê bikar anîn dimethyl fumarate ye. Tecfidera� (navê qeydkirî yê dimethyl fumarate ji hêla Biogen ve) bi bandor ji bo dermankirina formên vegerê yên skleroza pirjimar di hejmareke mezin a nexweşan de tê bikar anîn [152]. Lêbelê, bandorkeriya karanîna çalakkerên Nrf2 ji bo dermankirina nexweşiyên înflamatuar pêdivî bi erêkirina bêtir hewce dike da ku ji bandorên zirardar ên Nrf2 dûr bixe. Ji ber vê yekê, pêşkeftina dermankirinên ji bo çalakiya dijî-iltîhaba ku ji hêla Nrf2 ve hatî navbeynkirin dikare bandorek klînîkî ya girîng hebe. Lêkolînên domdar ên rêça sînyala Nrf2 li çaraliyê cîhanê ji bo pêşxistina ajanên dermanker ên pir-armanc ji bo kontrolkirina nîşanên iltîhabayê, û pêşîgirtin û dermankirina kanserê û her weha neurodegenerative û nexweşiyên din ên sereke têne veqetandin.
Di encamê de, Nrf2 di asta oksîterê de li ser laşê mirovan û hêdî dibe ku rêbazkirina antîxidant û enzyme û genên destnîşan dike. Ji ber sedemên zordarî ji ber zêdebûna asta oxidatîf ve bi nexweşiyên neurodegenerative ve girêdayî ye, Nrf2 dikare roleke girîng be di dermankirina pirsgirêkên tenduristiyê yên mîna nexweşiya Alzheimer de, di nav yên din de. Qada agahdariya me bi pirsgirêkên tenduristiya chiropractîk û spinal ve sînorkirî ye. Ji bo nîqaşkirina mijarê, ji kerema xwe ji Dr. Jimenez bipirsin an bi me re têkilî daynin915-850-0900.
Pirsgirêk Pirsgirêk: Pevçûnê Gelek Kuştina bêyî Surgery
Painşa çokan semptomek navdar e ku dikare ji ber cûrbecûr birîndarbûn û / an şert û mercên çokan çêbibe, tê de birînên werziş. Çok di laşê mirov de yek ji hevgirêkên herî tevlihev e ji ber ku ji hevberdana çar hestî, çar lîgament, tendonên cihêreng, du menisci û kartilan pêk tê. Li gorî Akademiya Amerîkî ya Bijîjkên Malbatê, sedemên herî gelemperî yên êşa çokê di nav de binavbûna patellar, tendinitis patellar an çokê jumper, û nexweşiya Osgood-Schlatter hene. Her çend êşa çokê bi îhtîmalek mezin di mirovên ji 60 salî mezintir de çêbibe jî, êşa çok dikare di zarok û ciwanan de jî çêbibe. Êşa çokê li gorî rêbazên RICE dikare li malê were derman kirin, lêbelê, birînên giran ên çokê dibe ku guhdana bijîjkî ya tavilê hewce bike, tevî lênihêrîna chiropractîk. �
Nexweşiyên neurodegenerative, wekî nexweşiya Alzheimer û nexweşiya Parkinsonê, bi milyon kesan re bandor dike. Celebên alternatîfên cûda hene ku ji bo nîşanên gelek nexweşî yên neurodegenerative derman bikin, lêbelê encam pir caran sînor dike. Lêkolînan lêkolînên lêkolînan da ku sedema zextên oksîterî ji aliyê faktorên hundur û derveyî ve dibe sedema sedemên ji bo nexweşiyên neurodegenerative dibe. Ew faktoriya veguherînê, Nrf2, biryar da ku wekî mechanîsyona parastina sereke li dijî tedawiya oxaxative. Armanca jêrîn li jêr e ku bandorên xuya dike Nrf2 li ser nexweşiyên neurodegenerative.
Modulatoriya Proteostasis ji hêla NRF2 veguhestin
Nexweşên neurodegenerative bi kombûna komîteyên taybetî yên taybet re girêdayî ye, pêwendiya hevpeymaniyê di navbera mîzê û mirinê yên proteotasis de pêşniyar dikin. Proteostasis ji bo hemû pêvajoyên veguhestin ku hucreyên pirrjimar û pirzimanî ji bo torê ya berfireh dike ku kontrola rêyên rêwîtiyên gene, gene expression û protein degradasyonê ve girêdayî ye. Vê çavkaniyê hewldanên hewceyê ku di binpêkirina transîtalîzmê ya prototasis de ji hêla NRF2 (nimûneya nukleer (erythroid-derived 2) -ek 2-ê veguhastin veguhastin. NRF2, wekî rêbazek masterê wekî bersivê hucreya antioxidant tête dîtin, tevî ku ew niha wekî komeleke sereke ya pîşesaziya veguherînê ya ku prototasis diparêze. Wê ku em ê gotûbêj bikin, NRF2 dikare wekî hub-ê ku têguhestinên acîl ên ku ji berhevkirina protein-nexşedî ji bo ku ji bo bersivê veguherînek veguherînek hevdeng û dravîlekî çêbikin. Ev xebatên bi NRF2 re têkildarî girêdayî cinsên cinsî yên di fîzîkî ya fîzolojî, proteasome û autophagy ve girêdayî ya endoglasmîk a tevlêbûnê pêk tê.
Faktorê Nukleer (erythroid-derived 2) -like 2 (NRF2), proteinek bingehîn a leucine-zipper e ku îro îro wekî rêbazek masterê navmalê malostasî tê de tête dîtin. Ew gotina zencîra 250 ya ku bi gelemperî cis-acting enhancer ve girêdayî ye kontrola elementên antioxidant (RED) [1], [2], [3], [4], [5] binpêkirin. Ev genes di çarçoveya I-II, II û III re reaksiyonên teqokrasiyê, glutathione û metokolîzma peroxedoxîn, hilberîna NADPH bi rêya pisporê fosphate û malz enzyme, oxaxeniya fatiyayê, metabolîziya asûmet û proteotasis [6] beşdar dibin. Bi karên van cytoprotîkî ve tê dîtin, ew dikare dibe ku yek ji NRF2-ê dermanekolojîk ya ku bandorên sereke yên sereke yên kevn ên kêmtir dibe, kêmtir dibe, tevlî xweseriya germî, germ û proteotxîk. Roja NRF2 di modulasyonê ya parastina antioxidant û çareserkirina hesamiyê de gelek lêkolînên di mijarên [7] de hate binçavkirin. Li vir, em ê li ser prototasis in, rola navxweyî ya kontrola protein, dabeşkirinê, bazirganî û hilweşandinê li ser rola xwe bikin. Wek mînak dê di çarçoveya nexweşiyên neurodegenerative de têne dayîn.
Nîşanek gelemperî ya nexweşiyên neurodegenerative rûdana kombûna bêserûber a hin proteînan e. Ji ber vê yekê, komikên proteîn ên çewtî yên? -Sînukleîn (? -SYN) di nexweşiya Parkinson (PD), plaqe? -Amîloid (A?) Û tevnegirên neurofibrilarî yên TAU yên hyper-fosforîlekirî de hene di nexweşiya Alzheimer (AD) de, nêçîrîn (Htt) li Nexweşiya Huntington (HD), superoksîd dismutaza 1 (SOD1) û TAR DNA girêdana proteîna 43 (TDP-43) di skleroza lateralî ya amiyotrofîk (ALS), proteîna prîonê (PrP) di encephalopathiyên spongiform, û hwd. Bi tevahî proteîn dikarin bandorê li çendan bikin riyên hucreyî, ku di encam de dibe ku bandor li ser ast û çalakiya NRF2 bike.
Cenazeyên Cûda yên Qanûna Cûreyên Çalakiya NRF2 Kurt
Di bin mercên fizîkî de, hucreyên asta NRF2 kêmtir ji ber veguhertina rasterast. Bi bersivên cuda yên cûda, NRF2 protein tê hesibandin, navberî navçe û veguhastina jinan. Ji ber vê yekê, birêvebirina asta protein ya NRF2 e ku girîng e ku divê divê nîşanên pêwendiyên erênî û neyînî pêk bînin. Wekî ku em ê dê bêtir gotûbêj bikin, NRF2 bi rêya mekanîzmayên celebên pirrjimar ên bi awayekî rasterast û bandorek ve çêtir dibe, lê ji aliyê din ve jî NRF2 dikare di pêvajoya duyemîn de, da ku ji bo bersiva wê veguherîne.
Ji dîtina kevneşopî, çalakkirina NRF2 wekî encama bersiva şaneyê ya li hember oksîdan an sazûmanên elektrofîlîk hate hesibandin. Di vî warî de, adapterê ubiquitin E3-lîgaze Kelch-like ECH-related protein 1 (KEAP1) roleke girîng dilîze. Dê hûrguliyên molekulî di Beşa 4.1 de bêtir bêne vegotin. Bi kurtahî, KEAP1 ji ber bermahiyên krîsteîn ên krîtîk ên ber bi ubiquitination û hilweşîna proteasomal a NRF2 ve wekî sensorek redox tevdigere. Ji bilî vê modulasyona klasîk, NRF2 bi bûyerên îşaretê bi rengek kûr tê tertîb kirin. Bi rastî, kînazên cihêreng hatine fosforîlekirin û nîzamkirina NRF2. Mînakî, NRF2 dikare ji hêla mitogjen proteîna kînazên aktîfkirî (MAPK) fosforîlasyon bibe, her çend tevkariya wê di çalakiya NRF2 de nediyar bimîne [8], [9], [10], [11]. PKA kînaz û her weha hin îzozîmên PKC [12], CK2 [13] an Fyn [14] fosforîlata NRF2 aramiya wê diguherînin. Xebata berê ya ji koma me ragihand ku glîkojen sentez kinse-3? (GSK-3?) Bi veqetandina nukleerê û hilweşîna proteasomal NRF2 asteng dike [15], [25], [26], [27], [28], [29], [30]. Dê hûrguliyên molekulî di Beşa 4.1 de werin nîqaş kirin. Wekî din, NRF2 ji celebên din ên rêziknameyê re tê şandin. Mînakî, acetîlasyona NRF2 ji hêla CBP / p300 çalakiya xwe zêde dike [17], dema ku ew ji hêla miR153, miR27a, miR142-5p, û miR144 [16], an jî ji hêla metîlasyona giravên sîtosîn-guanîn (CG) ve di nav pêşvebirê NRF2 de tê asteng kirin [18]
Impact of Protein Aggregates on Mechanism Regulatory NRF2
Di vê beşê de em ê li ser çawa ku çawa proteînkirina şaşên neheqandî dikare dikare bandora NRF2 bandora hin rêyên ku ji jorên jorîn re wek mînak wek illustratorî pêşkêş dikin pêşkêş dikin. Berî me, divê em bifikirin ku qezenckirina protein bi zirarê oxuxative ve girêdayî ye. Bêguman, danûstandinên danûstandinê û tevlihevkirina neheqetiya hilberîna hilberîna nermîner ên oxygenê (ROS) ji ji mitokondraya û çavkaniyên din [19]. Wekî ku jor gotiye, ROS dê li ser KEAP1 ya redox-hest-guhartinên ku ji bo serbestberdan, stabilîzasyon û niştecîhkirina nukleer ya NRF2 ve biguherînin.
Di derheqê proteînopatî de, mînakek bûyerên îşaretê yên bêserûber ku dibe ku bandorê li NRF2 bike ji hêla hîperaktîfkirina GSK-3 ve tê peyda kirin? di mîladî de. GSK-3 ?, ku wekî TAU kinase jî tê zanîn, beşdarî fosforîlasyona vê proteîna girêdayî mîkrotubulê dibe, di encama kombûna wê de, çêbûna tanglesên neurofibrillary û qutbûna veguhastina axonal (di [20] de tê nirxandin). Li aliyê din, GSK-3? asta û çalakiya NRF2 ku li jor hate behs kirin bi rengekî berbiçav kêm dike. Her çend bi gelemperî neyê pejirandin jî, cascade amîloîd pêşniyar dike ku A jehrîn e? olîgomer GSK-3 zêde dikin? çalakiya bi hevra TAU hîper-fosforîlasyon û mirina neuron [21], [22]. Modelên cihêreng hene ku vebêjin çawa A? GSK3- hez dike? çalakî. Mînakî, A? bi receptorê însulînê ve girêdide û rêyên îşaretkirina PI3K û AKT asteng dike, ku ji bo domandina GSK-3 girîng in? ji hêla fosforîlasyonê ve li paşmayîna N-termînalê Ser9-ê neçalak [23]. Li aliyê din, A-ya derveyî hucreyî? bi receptorên Frizzled re têkiliyê datîne, îşaretkirina WNT [24] asteng dike û dîsa di encama serbestberdana GSK-3 çalak de pêk tê ?. Bi kurtahî, A? kombûn dibe sedema hîperaktîfkirina anormal a GSK-3?, bi vî rengî bersivek guncan a NRF2 xerab dike.
Wekî ku di beşa jêrîn de nîqaş kirin, protein neheqandin çalakiya PERK û MAPK, ku bi riya NRF2 [31], [8], [9], [10], [11]. Wekî din, çalakiya CBP / p300 di gelek proteinopathies de [xNUMX] û kêmbûna globalbûna di DNA-Methylasyonê de di navên AD-ê de [32] jî nîşan dide, ji ber vê yekê bingehîn pêşkêş dikin ku ji bo van peywendiyên NRF33 di encamên têkildariyê de bibînin.
Me û yên din di nav neropên PD û AD-ê de zêdebûna di asta protein ya NRF2 û hin armancên wê, wekî oxygenase hema 1 (HMOX1), NADPH quinone oxidase 1 (NQO1), p62, etc., hem jî ji aliyê immunoblot û ji hêla immunohistochemistry [34], [35], [36], [37], [38], [39]. Li ser van ramanên NRF2 li ser van nexweşiyên wekî mizgîniya mêjî ya ku ji bo nirxên homostatîk vedigerin tê şirove kirin. Lêbelê, lêkolînek din jî diyar kir ku NRF2 piraniya cytoplasm a nebronên Hippocampal ên herêmî ye, pêşniyar kir ku çalakiya NRF2 di mêjûyê de [40] kêm kir. Ew têgihîştin ku cudahiyê van van çavdêriyan di guherînên fikrên têkildarî NRF2 de di çarçoveyên pêşveçûna neurodegenerasyonê de kontrol dikin.
Sîstemên sereke yên sê-sêyan bi proteotasisan re, bi navê bersiva protein (UPR), sîstema proteinome (UPS) û autophagy. Bêguman, em delîlên nirxandina NRF2-ê wekî hub-nîşeyên acîl ên ku ji hêla protein-êkûterên bi kompaniyên proteîn ên derveyî ve çalak ve girêdayî ye.
NRF2 Tevlêbûna Pirsgirêka Proteen (UPR)
NRF2 Çalakiya Endamê ya UPR
Di ER de qatbûna proteîna oksîdatîf ji hêla gelek rêyên diyar ve tê ajotin, ya ku herî tê parastin proteîna disulfîd-îzomerazê (PDI) û sulfhîdrîl oxîdaza endoplazmî ya oksîdoreductin 1 (ERO1? Û ERO1? Di memikan de) wekî donorê dîsulfîdê digire nav xwe. Bi kurtahî, PDI ji ber kêmkirin û oksîdasyona amînoacîdên xweya cîsteîn, çêbûn û şikestina bendên disulfîdê yên di navbera bermahiyên cysteine de di nav proteînan de katalîzekir, PDI ji hêla çalakiya enzima xaniyê malê ERO1 ve tête nûve kirin, ku girêdanên dîsulfîdê ji nû ve vedigire nav PDI [41]. Oksîjena molekulî qebûlkerê elektronê ya termînalê ya ERO1 e, ku ji bo her girêdana disulfîdê hilberîn mîqdarên stoçîometrîkî ya hîdrojen peroksîdê çêdike [42]. Peroxidases (PRX4) û glutathione peroxidases (GPX7 û GPX8) enzîmên sereke ne ku hîdrojen peroksît di ER de kêm dikin. Dema ku ev pergala oksîdo-kêmker bi rêkûpêk neyê xebitandin, berhevkariya anormal a proteînên çewt pêlkirî di ER de çêdibe û komek îşaretên bi navê bersiva proteîna nevekirî (UPR) ji sîtoplazmayê û navikê re tê veguhestin da ku homeostasiya ER ji nû ve were sazkirin [43]. Sê proteînên têkildar ên membran ji bo hesandina stresa ER li eukaryotiyan hatine destnîşankirin: faktorê veguheztina 6 çalakkirin (ATF6), ER eIF2 ya pankreasê? kinase (PERK, di heman demê de proteîna kinase-ER-kinase-wekî-çalak RNA-aktîfkirî), û kinase1-hewceyî înositol (IRE1). Domîneya lumînal a her sansorek girêdayî ye ku bi chaperona 78 kDa re tê gotin ku proteîna bi glîkoz-rêkûpêk tê binavkirin (GRP78 / BIP). BIP li ser stresa ER ji hev vediqete ku proteînên neqişandî bi hev ve girêbide, dibe sedema çalakkirina sê sensoran [44].
NRF2 û NRF1 her homolog, herweha bi bersiva antioxidant re, beşdarî veguherandina UPR bi nucleus beşdarî beşdarî. Di rewşeke NRF1 de, ev proteîn li li membrana ER-ê ye û di nav veguhertina nukleerî ya nukleer a deglycosylation an jî paqij. Piştre, çalakiya UPR tête pêvajoyê ya NRF1 û kombûna nukleer ya ku di encama nukleerî ya nukleerî de pêk tê. Lê belê, kapasîteya veguherandina veguherketin ARE-jinan yên vê NRF1 jî hîn jî di bin gotûbêjê de [45] ye.
Glover-Cutter û hevserokên karkerên çalakiya NRF2 anthologue C. C. elegans, SKN-1, bi ER-strên cuda cuda nîşanî. Xwenîşandanek SKN-1 zêdebû li ser navendên UPR-ê girêdayî bû, bi including IRE1 an ortologues-PERK-ê de [46]. Di hucreyên perk-kêm de, têgehek protein-nehêleya nehêle dibe sedema kombûnên endogenous peroxîd û paşê apoptosê [47]. Effector ji hêla PERK ve tê bikaranîn ER ji ji van peroxoxên ERF2, ji ber ku ev rapor kirin ku NRF2-NRF40-PER1 PERK PER31, bi vî awayî wê ji aliyê KEAP1 ve veguhestin [2]. Têkilî ya ASK1 jî di vê rêgezê de bi rêya vê rêwîtiya TRAF48-kartê çalakiya IRE2 [1]. Tevî ku rola MAPKs di rêznameya NRF2 de hîn jî nexşandî ye, ev yek dihat pêşniyar kirin ku dibe ku rêvekirina IRE1-TRAF2-ASK49-JNK NRF1 çalak bike [1] (Xigê 1). Bi balkêş e, di nav. Cîhanên elegans û mirovên mirovan, pêşniyar dike ku cinazeya sulfenîlasyonê kinase ya IRE38-ê li ser çalakiya wê ya UPR-IRE2-mediated tête dike û bersiva antoxxantant p1 bi NRF38 ve tête destpêkirin. Daneyên daneyên ku IRE2 çalakiyek kevneşopî ye ku wekî cytoplasmic sendinel ku çalak dike p50 û NRFXNUMX [XNUMX].
Qanûna 1 Qanûna NRF2 ji aliyê UPR. Gelek pro proteins di hundurê retokumasyona endoplasmîk de dikare pêvajoya bersivên hilberîn (UPR) dest pê dike. Ya yekem, Baperona BIP ji ji navenda navendî ya ER-ê ve hat serbest kirin IRE1 û PERK ji bo proteînên nehêl / misfolded. Ew dîmerîzasyon û trans-auto-phosphorylasyonên domên cytosolic ên xwe dike. Encamên çalakiya PERK-ê li rasterast NRF2 li Ser40, ku ji bo NRF2 veguhestina nucleus û çalakvanên armancên hedef. Activation of IRE1 veguhastina TRAF2 veguhestin û piştî çalakiya ASK1 û JNK phosphorylation. Wekî ku JNK ji bo phosphorylate ve hat ragihandin û NRF2 çalak bike, ew e ku fikir e ku ji bo çalakiya IRE1 dê çalakiya NRF2 zêde bibe.
Li ser înfeksiyona UPR-ê gelek lêkolîn bi hilgirîna proteîn glîkozîlasyon tunîkamîsîn re hatine kirin. NRF2 ji bo pêşîlêgirtina mirina şaneya apoptotîk a ku ji hêla tunicamycin ve hatî çêkirin girîng xuya dike [31] û çalakkirina wê di bin van mercan de ji hêla hilweşîna otofagî ya KEAP1 ve tê rêve kirin [51]. Li gorî vê yekê, bêdengkirina shRNA-navbeynkariya derbirîna NRF2 di şaneyên? TC-6 de, rêzikek însulînoma murî, bi awayekî girîng sîtotoksîksiyona ku ji hêla tunicamycin ve hatî çêkirin zêde kir û bû sedema zêdebûna derbirîna nîşana stresê ya ER-apoptotîk CHOP10. Li aliyê din, çalakkirina NRF2 ji hêla 1,2-dithiole-3-thione (D3T) sîtotoksîksiyona tunîkamycîn kêm kir û vegotina CHOP10 û PERK lawaz kir [52]. Bi balkêşî, neuronên bîhnxweş ên ji bo sepandina pergalê ya tunicamycin radest kirin NRF2 li rex endamên UPR-endamên din ên wekî CHOP, BIP, XBP1 zêde kir [53]. Van encaman ji bo lêkolînên vivo hatine dirêj kirin, ji ber ku enfeksiyona devokî ya tenurîn a tunicamycin di maran de derbirîna PERK û NRF2 di hîpokampus de digel kêmasiyên zanistî yên girîng, fosforîlasyona TAU û depozîtanên A? 42 zêde kirin [54].
NRF2 Ji bo Serastkirina ER Fîzolojiya Key Genes Up-Regulates
Ji bo domandina kîmyaya disulfîd pêdivî ye ku lumena ER ji sîtosolê bi dabînek pirr GSH hebe. NRF2 enzîmên girîng ên metabolîzma GSH di mêjî de module dike, wekî veguhastina cystine / glutamate,? -Glutamate cysteine synthetase (? -GS), glutamate-cysteine ligase katalîtîk û yekîneyên moderator (GCLC û GCLM), glutathione reduktase (GR) û glutathione peroxidase (GPX) (li [55] hatî nirxandin). Têkiliya NRF2 di parastina GSH de li ER ve tête piştgirî kirin ku çalakkirina dermankolojîk an genetîkî ya NRF2 bi zêdebûna senteza GSH bi GCLC / GCLM encam dide, dema ku astengkirina vegotina van enzîmên bi NRF2-xistinê dibe sedema kombûna zirarê proteînên di nav ER de dibe sedema çalakkirina UPR [56].
Di C. elegans de gelek pêkhateyên genên UPR hedef ku ji hêla SKN-1 ve têne birêve birin, Ire1, Xbp1 û Atf6 jî hene. Her çend NRF2 di memikan de vegotina çend genên peroksîdaz (PRX) û glutathione peroxidase (GPX) bi rêkûpêk dike (di [57] de hatî nirxandin), tenê GPX8 enzîma ER-herêmî ya dilsoz e, nîşana vegera KDEL vedigire [58]. Windabûna GPX8 dibe sedema çalakkirina UPR, herikîna ERO1? -Piroksîjena hîdrojenê ber bi sîtosol û mirina şaneyê ve. Hîdrojen peroksîda ku ji ERO1 hatî ye? çalakî ji ber çalakiya lihevhatî ya GPX8 û PRX4 nikare ji ER ber sîtosol belav bibe [59]. Di vî warî de, analîzek ji array derbirîna rêça-genê parastina antioxidant bi karanîna RNA ji celebê çolê û tevna mişkên NRF2-nal, diyar kir ku vegotina GPX8 di nebûna NRF2 de kêm-rêkûpêkkirî bû [60]. Li gorî vê yekê, analîzên transkriptomê ji nimûneyên nexweşan ên ku ji neoplazmayên mîeloprolîferatif, polycythemia an myelofibrosis êş dikişînin, nexweşî bi stresa oksîdatîf û iltîhaba kronîk a pola nizm re têkildar in, asta vegotina her du NRF2 û GPX8 bi mijarên kontrolê re nîşan dide [61]. Hîn lêkolînên ku bi taybetî GPX8 di parastina mejiyê mirovan de hene tune ne lê vekolînek transcriptome di mişkan de zêdebûna GPX8 ya telafîker di bersivdana toksîna Parkinsonian MPTP de nîşan dide [62].
Impact of NRF2 on Dysregulation li Nexweşên Neurodegenerative
Malfunction of PDI enzymes û çalakiya kronîk ya UPR dikare piştre neurodegenerasyon dest pê bike yan zûtir bike. Nexweşên nexweşî-neurons, modela heywanên nexweşî neurodegenerative û herweha wekî tissuesên paşîn ên mirinê yên paşîn-mêrem li ser piraniya van tengahiyên li ser gelek rêzikên UPR-marker diyar kirin. Guhertina pîşesaziya PDI / Rêveberiya UPR ya di nexweşiyên neurodegenerative vekirî de di binçavkirin [63] ve hatî binçavkirin, lê jêrîn li ser hûrgelên paşîn ên morgê mêjî bêne bêne kirin. Pîvanên PDI di neronên tengalê de têne zêdekirin û li Lîreyên AD û PD nexweşan, bi [64], [65]. PDI û ERP57 li CSF ji nexweşiyên ALS û di nav mirên CJD de [66], [67], [68] ve têne rêve kirin. BIP, PERK, IRE1 û ATF6 di navnîşanên ji AD, PD û ALS de [69], [70], [71], [67] têne çêkirin. BIP, CHOP û XBP1 di binpêkirinên mêjûya mîzê de ji HD [72], [73] ve têne bilind kirin. Ji bilî, verastkirina uplêdana ERP57, GRP94 û BIP di nexweşiyên cortex ji CJD nexweşan de [74] hate dîtin. Bi tevahî, ev delîl diyar dike ku ji berhevkirina hilberên derfetên ku di mêjû parenchyma de çalakiya xurt û çalakiya demokrasiyê ya UPR de dibe. Bi balkêş e, lêkolînek dawînek heye ku çalakiya NRF2 bi PERK di destpêka AD-ê de girêdayî ye. Di vê lêkolînê de, nivîskaran lêkolîn kir ku tedbîrên oxoxative guhertinên di NRF2 de guhertin û UPR dikare dibe ku di rêgezên AD-ê de ji hucreyên xwînê yên peripheral û nimûne pergala mîkrojenên mîkrojen ên trajnîk ên di asta nexweşî de pêk tê. Stressed oksîdative û pSer40-NRF2 zêdebûna hucreyên mononolojî yên xweserî yên mirovî yên ku bi astengiya cognitive nefretê vekirî vekirî hate dîtin. Ji bilî wan, ew li ser astengên ER calcium malostasis û li ser van hucreyên ER-stressî yên ku bi astengiya cognitive nefret û nerm e-nerm [75] rapor kirin rapor kirin.
Rêziknameya Hevpar a NRF2 û Pergala Ubiquitin Proteasome (UPS)
Nûnerên UPS-ê yên NRF2-Protein
UPS beşdarî hilweşandina xerît û neheqên şaş û tevlîhevkirina asta kolektîfên sereke di cytosol û nucleus de beşdar dike. Navenda navendî ya vê pergala enzyme pir mezin e ku bi navê 20S veguherînek protokytîk e. 20S hilberên proteinsome dezgehên hilberîneyên proteîn vekirî, lê ji bendên bi proteîneyên protein yên rêbazên cuda yên taybetmendî û çalakiya xwe veguherînin biguherînin. Ji bo nimûne, her wiha ji du beşên 19S yên ku ji bingeha 20S re proteasome 26S pêk tê dike û her taybetmendiya li ser protein [76], [77] biguherînin. Pêdiviya Proteasomal hewceyê bêdengkirina covalent ya ubiquitin. Conjugation of ubiquitin through a step-by-step mechanismism cascade. Ya yekem, ubiquitin-activating enzyme E1 çalakiya ubiquitin di an ATP-reaksiyonê hewce dike. Piştre, an E2 enzyme (protein ubiquitin-carrier protein an ubiquitin-conjugating) veguherandina ubiquitin ji E1 ve li substrate veguherîne ku bi taybetî bi endamê endamê malbata ubiquitin-protein bi navê E3 ve girêdayî ye. Tevî ku çarçoveyek rastîn ê protein-ê di çarçoveya xwezayî ya ubiquitin de girêdayî ye, ev pêvajoyê bi gelemperî di hilweşandina 26S de [78] encam dike.
E3-ligase KEAP1 herî qedexekirî ya NRF2 e. Mîkrofîzma ji biryara KEAP1 diyar eşkere dike ku çiqas asta NRF2 bi lezgehên oxîdant re bigire. Di bin şertên basal de, NRF2-yê KEAP1-ê ji hêla homodîmer-ê KEAP2 ve tê veguhastin, ku yek naneyên NRF1 bi du amino-anilên bi low (aspartate, leucine, glycine) û bilind (glutamate, threonine, glycine, glutamate; ETGE) girêdayî. Têkiliya bi KEAP2 re peyda dike ku NRF3 ji pelê proteînkirina CULLIN1 / RBX1-ê, ku di encama nerast û paşê proteasomal de paşde de paşerojê pêşkêş dikin. Lêbelê, guhertina redox KEAP2 pêşkêşkirina NRF3 ya UPS bi CULLIN1 / RBX2 ve tê nîşandan. Wekî encamek, NRF1 nû hate revandin KEAP79-dependent dezgehkirin, di nav nucleus de tête kirin û çalakvanên hene. Jinan [80], [81], [82], [XNUMX].
Adapter? -TrCP E3-ligase di heman demê de homodîmer e ku beşdarî bûyerên îşaretê yên têkildarî fosforîlasyona NRF2 ji hêla GSK-3? Ve dibe. Ev kinase bermahiyên serîn ên NRF2 fosforîlate dike (aspartate, serîn, glycine, isoleucine serine; DSGIS) da ku qadek hilweşînê çêbike ku paşê ji hêla? -TrCP ve tê nas kirin û ji hêla proteîna komeleyê ve ji hêla tevliheviyek CULLIN1 / RBX1 ve tête nîşankirin. Nasnameya asîdên amînoyî yên taybetî ku ji hêla GSK-3 ve têne fosforîlekirin? di vê degronê de ji hêla mutagneziya malper-rêvekirî ya qada Neh6, elektroforeza 2D-gel [15], [26] û spektroskopiya girseyî [83] hate pêk anîn. Di encamê de, astengkirina GSK-3? ji hêla dermanên pir bijarte an siRNA ve li dijî îzoformên GSK-3 di zêdebûna asta proteîna NRF2 de derket. Encamên manend ên bi siRNA re li dijî îzoformên 1-2. a? -TrCP hatin dîtin. Stabilîzasyona NRF2 li pey GSK-3? astengkirin di fibroblastên embriyoyê mişk-kêmasiyê KEAP1 û di mutanta jêbirinê ya NRF2 ya ektopîkî ve hatî îfadekirin de ji bermahiyên ETGE krîtîk ên ji bo girêdana pêwendiya bilind a KEAP1 ve tune, çêbû, bêtir rêziknameya serbixwe ya KEAP1 nîşan dide.
Di çarçoveya nexweşiyên nerjenegerîner de, em dikarin modulasyona NRF2 ji hêla UPS-ê ve bi du awayên cûda vebînin. Li aliyek, pergala KEAP1 dê hevsengiya redoxê ya ku ji berhevoka proteîna çewt hatî hilberandin hîs bike, dema ku GSK-3 /? - Axa TrCP dê wekî beşdarvanek çalak be di îşaretkirina veguhastinê de ku bi windabûna proteostasis ve hatî guhertin (Hêjîrê. 2).
2 Tiştek UPS bi kurtî asta NRF2 kontrol dike. Di bin mercên homeostatîkî de, asta NRF2 kêm bi çalakiya adapterên E3-lîgases KEAP1 û? -TrCP têne domandin. Çep, NRF2 bi navgîniya motîfên hevalbendiya kêm (DLG) û bilind (ETGE) ve bi warên Kelch ên homodimer a KEAP1 ve girêdide. Bi navgîniya qada BTB, KEAP1 di heman demê de bi kompleksa CULLIN3 / RBX1 ve tê girêdan, ji hêla 2 S proteasome ve bikarneanîn û xerakirina NRF26. Wekî din, GSK-3? fosforîletên Ser335 û Ser338 bermayiyên NRF2-ê çêbikin ku qada hilweşandinê (DpSGIpSL) biafirînin ku dûv re ji hêla adapterê ubiquitin ligase? -TrCP ve tê pejirandin û ji hêla texrîbata proteasome ve ji hêla tevliheviyek CULLIN3 / RBX1 ve tête nîşankirin. Rast, Bi rûdana cinsên oksîjena reaktîf an elektrofîl re bermahiyên Cys ên krîtîk di KEAP1 de têne guhertin, dibe ku KEAP1 nikaribe bi NRF2 an CULLIN3 / RBX1 re têkiliyek bi bandor bike û dûv re jî ev faktorê veguhastinê çalakiya nîv-jiyan û veguheztina xwe ya ber bi ARE-genan ve zêde dike. Rêyên îşaretê yên ku dibin sedema astengkirina GSK-3 ?, fosforîlasyona AKT-a wusa li Ser9, di encama NRF2-ê de ji hêla proteasome, kombûn û derxistina genên hedef ve xera dibe.
NRF2 Bi Tevgera Transcriptional ya Proteasome Subunits Bi UPS Çalakiyê zêde dike
NRF2 îfadekirina çend binavên proteasomê bi rêkûpêk dike, û bi vî rengî şaneyê ji berhevkirina proteînên jehrîn diparêze. Bîst genên têkildar proteasome û ubiquitination xuya dike ku ji hêla NRF2 ve têne rêkûpêk kirin, li gorî analîzek mîkro-fireh a fireh ji RNA kezebê ku bi D2T induktorê NRF3 ve hatî saz kirin [84]. Di lêkolînek paşîn de, heman nivîskaran diyar kir ku vegotina piraniya bin-yekên 26S proteasome di mişkên bi D3T hatine dermankirin de di kezebê de sê qat zêde bûne. Asta proteîna subunit û çalakiya proteasome bi hevrêzî zêde bûn. Lêbelê, di mişkên ku faktora transkrîpsiyonê NRF2 têkçûyî de hîç indukasyonek nehate dîtin. Çalakiya danasîner a bin-yeka proteasome ya PSMB5 (20S) bi zêde îfadekirina NRF2 an dermankirina bi çalakvanan re di fibroblastên embrîyonîk ên mişk de zêde bû, û ARE di danasînerê proximal ê PSMB5 de hatin diyar kirin [85]. Çalakkirina dermankolojîk a NRF2 di asta vegotinê ya yekîneyên proteasome yên nûner (PSMA3, PSMA6, PSMB1 û PSMB5) de tenê di fibroblastên mirovî yên ne-pîrbûyî de NRF2 ya fonksiyonel heye [86]. Çalakkirina NRF2 di dema adaptasyonê de ji bo stresa oksîdatîf di vegotina bilind a PSMB1 (20S) û PA28 de encam dide? subunits (an S11, regulator proteasome) [87]. Wekî din, encamên ji şaneyên stem ên embrîyonî derket holê ku NRF2 îfadekirina proteîna gihîştina proteasomê (POMP), çeperek proteasome kontrol dike, ku di encamê de pirbûna xweser-nûvekirina hucreyên rehên embrîyonîk ên mirov, sê ciyawaziya tebeqeya germê û ji nû ve bernamekirina hucreyê modûl dike [ 88] Hemî bi hev re, van lêkolînan diyar dikin ku NRF2 vegotina pêkhateyên sereke yên UPS-ê bi rêkûpêk dike û ji ber vê yekê bi rengek çalak beşdarî paqijkirina proteînên ku wekî din jehrîner dibin dibe.
Nexweşên Neurodegenerative Di NRF2-UPS Axis
Rola UPS di nav nexweşiyên neurodegenerative devere deverek xeletiyek pirfireh e. Di lêkolînên destpêkê de çalakiyên proteasom kêm kirine ku di nav çend nexweşiyên neurodegenerative yên nexweş de bandor kirin. Lêbelê, xebatên din ên li vîtro û karûbarên vivo nehatine guhertin an jî çalakiya proteosome jî zêde bûn (di nav [89] de têneçavkirin. Ji ber vê rêjeyê ji bo vêhevgirtina vê gengaziyê ev e ku ev asta UPS beşên pêşveçûnê di pêşveçûna nexweşî û di nav deverên mizgehên wekî ku ji bo NRF2-armanc hatine pêşniyar kirin.
Tevî vê nakokiyê, divê bêne xuyakirin ku hiqûqa jimara jînên proteinsome yên wê dê UPS bi hêza paqijkirina proteinên zikmakî di mêjî de hêzdar bikin. Bi rastî, betalkirina NRF1, modulatorê bersivê antioxidant jî, di hucreyên neuronal de dibe sedema çalakiya proteasom û neurodegenerasyon. Tecrûbeyên tecrûbeyên kapîtalîzmê yên Chromunusê û analîzkirina transcriptional nîşan da ku PSMB6 bi NRF1 ve têne birin. Herweha, profesyoneliya gene ya nîşankirina nasnameya NRF1 wekî nîjonê ya neuronsên veguhastina keyxebûra sereke, ku pêşniyar dike ku nerazîbûna NRF1 dikare bibe nexweşiyên neurodegenerative nexodegenerative. [90]. Bi balkêş e, NRF1 û TCF11 ya dirêj a ku navê hilberîna hilberên proteinsê li ser hilberên proteinsome li ser proteinsome protokolê nîşan da ku nîşana çalakiya proteolytîk kêm bike [91].
Li ser NRF2, têkiliyek di navbera kêmkirina NRF2, RPT6 (19 S) û PSMB5 (20 S) asta di navîn de DYA-DJ-1-kêşê ya mîkrobatê bi bi neurotoxin paraquat re tête kirin [93]. Ji bilî, kargeha xwezayî ya sulforaphane (SFN) dide ku wêneyê bêhtir NRF2 wekî modulatorek girîng a UPS. Di tecrûbeyên vitro yên bi neuroblastoma neuro2A de hucreyên nerazîbûna pêşveçûna behsa catalytîk ên proteasome û herweha jî çalakiyên peptidase di bersiva SFN de hate belav kirin. Ev derman hucreyan ji hîdrogen peroxide-cytotoxicity û oxygenê protein-ê bi awayekî girêdayî fonksiyonê proteinsome re girêdayî girêdayî parastin [94]. Herweha, Liu û karmendên karkerên aksiyonê kar kir ku ji bo bersiva SFN di mêjû de çalakiya UPS çalakiya kontrol bikin. Ev pişkên bi qewimî li protein (GFP) ji hêla veguhestina hilweşandina mûzayê vekirî diyar kir ku ji hêla UPS (GFPU) ve hilda xwe dikeve. Di cortexê cerebal de, SFN bi asta parallel a di chymotrypsin-like (PSMB5), caspase-like (PSMB2) û çalakiyên hewsîn-like (PSMB1) yên 20-S proteasome kêm kir. Ji bilî, tedawiya Huntington-SFN-ê vekirî diyar kir ku çalakiya NRF2 mHtt hilweşandin û mhtt cytotoxicity kêm bike [95]. Mîkrofîzma sereke ya SFN bi veguhestina NRF2 ve ye [96]. Beşdariya taybetî ya NRF2 divê sîstema NRF2-null di xebatên bêtir xebatên kar bikin.
Têkiliya Karûbarê ya Navbera NRF2 û Macroautophagy
NRF2 Protein Hilbijartin ji hêla Adaptter Protein P62 ve têne guhertin
Autophagy ji bo pergala cytosolîk di hundirê lîsosomes de veguhestin. Ev pêvajoyê ji bo paqijkirina dirêj-jiyan û protein-nefret û herweha organîzeyên zirarê tê bikaranîn. Girêdana yekser di navbera NRF2 û autophagy cara yekem di pêvajoya p62-adapter de, SQSTM1 jî [xNUMX], [97], [98], [99], [100] tê şandin. Ev protein şuttên ubiquitinated proteinsomal û makîsomal degradation machinseries and sequesters proteins in berî hilweşandina wan ji berhev kirin. P101 ji bo ubiquitin-girêdayî (UBA) domain, ji bo pêkanîna proteinên ubiquitandî, û herêma LC62-interacting (LIR) ji bo integrasyona autophagosomal membrana LF3 bi xweya autophagomomal.
Tevî ku pêwendiya p62-navbeynkariya NRF2 û genên wê yên armanc yekemcar di 2007-an de hate ragihandin [102], lê mekanîzmaya molekulî bi tevahî nehat fam kirin heya ku vedîtina têkiliya wê bi KEAP1 [103], [98], [99], [100 ], [101]. Komatsu û karkeran di p1-ê de KEAP62-a ku têkiliya KEAP1-ê (KIR) bi NRF2-ê re di heman bêrîka rûyê bingehîn de girêdide û bi hevalbendiyek girêdanê re dişibîne motîfa ETGE-ya di NRF2-ê de, pêşnîyar dike ku pêşbazî di navbera p62 û NRF2-ê de ye, nas kirin. Fosforîlasyona Ser351 di motifa KIR de di p62 (349-DPSTGE-354) de hate diyar kirin ku pêwendiya xwe ji bo KEAP1 zêde dike, bi girêdana NRF2 re dibe hevrik û destûr dide berhevkirina wê û çalakkirina transkrîpsiyonê ya genên armanc [98], [99]. Bi rastî, zêdeperestiya p62 bû sedem ku Ubiquitination NRF2 kêm bibe û di encamê de aram bibe û her weha genên wê yên armanc hedef were girtin [104]. Hin kînaz hatine pêşniyar kirin ku beşdarî fosforîlasyona p62 bibin. Armanca mammaliyan a tevliheviya rapamycin 1 (mTORC1) dibe ku têkildar be, lewra ku dermankirina bi ragîhandina mTOR rapamycin re fosforîlasyona p62 û xwarê-rêkûpêkkirina KEAP1 li ser dermankirina arsenît perçiqand. Di demên dawî de, hate xuyang kirin ku TGF -? - kînaza çalakkirî 1 (TAK1) dikare p62 jî fosforîle bike, hilweşandina KEAP1 û rêsazkirina NRF2-zêde bike. Nivîskarên vê lêkolînê pêşniyar dikin ku ev rêgezek ji bo birêkûpêkkirina redoxtasîzma hucreyî di bin mercên rewşa domdar de ye, ji ber ku TAK1-kêmasiya ROS-ê di nebûna tu oksîdanek biyanî de di tevnên cûda yên mişk de paralel bi kêmkirina asta proteîna NRF2 re-rêkûpêk dike [105 ]
A p62 ava kir ku hebûna UBA-ê jî hîn jî ji KEAP1 ve girêdayî ye, da ku ew têkilî li KEAP1 [101] ne girêdayî ye. Lêbelê, xerîbê p62 di melodografê Drosophila, navê Refê (2), nîvekek KIR nîne û nehêre bi DmKEAP1 re têkilî nake, lê teva ku ew bi rêya DBA-UBA DMKEAP1 ubiquitinated. Wekî din, DmKEAP1 dikare bi Directê bi Atg8 re têkilî bike (ji bo mammalian LC3 mumulî). Encamên KEAP1 di Atg8 û bixweberiya autophagy li ser NRF2 CncC û serbixwe li TFEB / MITF [106]. Têkiliya navbera NRF2 û autophagy xuya dibe ku her çiqas tête parastin, tevliheviya karûbarên wê dike.
Têkiliya NRF2 bi p62 encamek herdu kampanyona KEAP1 û hilweşandina KEAP1 di lysosome de. Bi xistina p62 bi siRNA bi dorpêçkirina KEAP1 di parallelê de kêmbûna NRF2 û cinsên armancên wê [101] dubare dubare. Di peymanê de, avakirina p62 gotina asta zêdebûna asta KEAP1 bi qaîdeyên şerr ên li dijî. Gelek têkildarî, zêdebûna di asta KEAP1 de ne ji hêla proteinsome protokolê ne bandor bû, lê di bin çavan-birêvebiriya autophagy kêm bûye [107]. Di rastiyê de, KEAP1 di hucreyên mammaliyan de di p62 û LC3 de [99], [100], [103] têne çêkirin. Hemî van daneyên pêşniyar dike ku KEAP1 pergala makrokotê ya macrootophagy e, lê ev pirsgirêk bêtir ji ber ku hebûna hebûna hin encamên nakokî tête analîzkirin. asta proteîna KEAP1 li mice Atg7-null [107] zêde bûn, a effector key yên macroautophagy [1], lê belê bêhêlin farmakolojîk ji macroautophagy bi torin64, E1 / pepstatin an bafilomycin bi ser neket ji bo berhevkirina KEAP107 [100],. Bi giştî, ev encam dide pêşniyar dikin ku p62 pevçûnên KEAP1 di navokên autofagîk de dibe û encama van encamên encamên KEAP1 devera autophagîk destûrê dide ku çalakiya NRF2 (Fig. 3). Du lêkolînên du şahidî ragihand ku sedsala sulfînîk kêm dike û SESTRINS di vê çarçoveyê de rola girîng girîng dike. SESTRIN 2 bi p62, KEAP1 û RBX1 re têkilî dike û p62-dependent degradation of KEAP1 û NRF2 çalakiya armancên target target [108]. Lêkolînek din nîşan dide ku SESTRIN 2 bi ULK1 û p62 re veguherand, fosphorylation of p62 li Ser403 ku ji bo KEAP1-109] proteînên kargo yên hêsantir dike.
Pîvanên 3 NRF2 P62 di pêvajoya adapterê de têne rêz kirin. Fosphorylation of Ser 351 in KIR motif of p62 (349-DPSTGE-354) bi mTORC1, TAK1 an jî kinên din ên din di encama zêdekirina encamên KEAP1 de ji ber ku wekheviya ETGE-ê di NRF2 de zêde dibe. Wekî encam, P62 dorpêçkirina NRF2 û KEAP1 bistîne. PIRN-LIR-P62-ê li LF3-ê li autophagosomal-membranê re têkiliyek dike, da ku puxînerê p62-KEAP1 di dawiya lîsosome de bête kirin. Wekî encamên NRF2 dikare bikar bîne, nucleus bi wergerandin û veguherandina belavkirina ya genê, p62. Ev mekanîzmîzma rêbazê bersiva NRF2 dakêşîne, wekî KEAP1 çalakiyek nû ye ku ji bo çalakiya NRF2 veguherîne.
Modulasyonê Genes of Macroautophagy by NRF2
NRF2 ji bo jenavên macroautophagy û herweha ew ji bo UPR û UPS tête dike. Pêşniyarên yekem ji xwendin bûn ku di p62-expression de hate diyarkirin ku li ser elektrofîlan, ROS û Oxîteya Nîtîk [110], [111], [112] tête kirin. Mîkrofîzma întegrasyonê çend salan paş bi vegotin ku dîtina p62 heye ku di xebata jene ya jene de [99]. Di lêkolîna dawîn de, deverên din yên xebatkar ên din jî hatin binçav kirin û pişta peymanên biyoformatîk û peymanên CHIP hatin dîtin. Ji bilî,, mîkrobeyên mîkroboks û neuronên cortical ji ji Nrf2-knockoutê nîşan da xuyakirin ku p62 expression, ku dikare bi NRF2-expressing lentivirus rizgar kirin. Bi vî awayî, kêmbûna NRF2 p62 asta di neuronên birîndar yên ji hippocampusê de [36] kêm kir. Ji ber vê yekê, pêşnîyar kir ku çalakiya NRF2 p62 pevçûnên zêde dibe, bi encamên KEAP1 hilweşandin û arîkariya bêtir NRF2 di loopê ya erênî yên erênî de. Vebijêrk ev ne-canonîk ya ya vegirtina NRF2 hewce dike ku di expressiona gene de guhertin û bersiva pêvajoya hucreyî bi demeke dirêj re têkildar be.
Parastina nasnameyê NDP52 hate nîşankirin ku ji aliyê NRF2 ve veguherandin. NDP52 di heman rengê de p62 dixebite, pêvajoyên ubiquitandî tê naskirin û bi LC3 bi rêya LIR-ê ve girêdayî ye, da ku ku cargoes di lîsosomes de neçar dibin. Niştecîhên pênc pêngavên di Ndp52 belavkerê DNA de hatin dîtin. Ji wan re ji hêla avakirina avahiyên mutant ên cuda yên ku ji bo NRF2-ya Ndp52-medyated-xpxNUMX-nexşebûn-an jî ji hêla CHIP-ê ve tê nas kirin. Têbigere, di nav hippocampus ya Nrf113-knockout de Ndp52 mRNA asta kêm bûn. Yek ji van herderan jî di bin lêkolînek serbixwe de wekî NRF2-RED-DEVE [2].
Lêbelê, rola NRF2 di modulasyona autophagyiyê de ne ku di pêvajoya van protein-nasnameyên van nasnameyê de ne sînor nîne. Ji bo ku bi rola NRF2 di modulasyonên xweser ên xweser ên xweser ên xweser ên xweser ên bêhtir de digerin, bi grûbê me yê ENCODE ji bo du proteînan, MAFK û BACH1, ku qeqên NRF2-rêvegirtin girêdayî. Bikaranîna pirtûkek ji jihevhatina JASPAR ve hatî çêkirin Li ser vê yekê, em ji gelek celebên xweser ên autofagagî ve hene. Diwanzdeh van van dîmenan de wekî deverên NRF2 di neh autophagy nine de, ku tê gotin ku di mîkrobeyên mîkroşê yên Nrf2-şikilê de kêmtir bû, lê dibe ku ji aliyê NRF2-lentivirus ve were veguherandin. Di xebata me de diyar kir ku NRF2 behsa behsa hin cinsên ku di pêvajoyên xweser ên autophagîk de tevlî çalak dike, tevlî pêvajoya autophagy (ULK1), têkoşîna otomatîk (p62 û NDP52), avakirina autofagosome (ATG4D, ATG7 û GABARAPL1), nerazîbûna (ATG2B û ATG5 ), û paqijkirina otolysosome (ATG4D). Di encam de, bersiva autophagy di bersiva hydrogen peroxide de bû ku dema NRF2 ne xuya bû [36].
Relevansiya NRF2-Navnîşên Macroautophagy Genesis Expression in Di nexweşiyên Neurodegenerative
Autofagy diyar kir ku di gelek nesrên neurodegenerative de rolek girîng e. [114] û derxistina xweseriya xweseriya neurodegenerasyonê li mûşê [115], [116]. Atg7-knockout icons revealed that encamên xweser ên autofagy di p62 kombûnên di ubiquitin-pêwendiyên erênî yên erênî de. KEAP1 di van saziyên tevlêbûnê de hate pêşîn kirin, ku bi rêberiya NRF2 û veguhestina cinsên armancên [103]. Girîng, pirhevkirina p62 bi hev re proteinên ubiquitinated di nasnameya neurodegenerative de, di nav AD, PD û ALS de [117] nas kirin. Di rastiyê de, neuronên bilind yên astengên APP û TAU yên nexweşan jî p62 û NRF2 ya nukleer diyar kirin, pêşniyar kir ku hewldanên wan hewldanên xweser ên înternetê yên xweseriya XXUMXê [36].
Kêmasiya NRF2 di çarçoveya AD de kombûna proteînê giran dike. Bi rastî, zêdebûna astên TAU-ya fosforîlasyon û sarkosîl-nayêne çareser kirin di mişkên Nrf2-knockout de tê dîtin, her çend cûdahî di çalakiyên kinase an fosfatazê de li beramberî paşnavê celeb-çolî tune be nayê dîtin [113]. Ya girîng, NDP52 hate nişandan ku bi TAU re di neuronên mûr de hev-lokalîzekirin û têkiliya rasterast di navbera fospho-TAU û NDP52 de ji hêla ceribandinên hev-immunoprecipitation ve hem di mişk û nimûneyên AD-ê de hate xuya kirin, û diyar kir ku ew rola wê di hilweşîna TAU de ye. Bi balkêş, bêdengkirina NDP52, p62 an NRF2 di neuronan de fosfo-TAU zêde kir [113], [118]. Wekî din, zêdebûnên APP-yên intraneuronal di hîpokampa mişkên APP / PS1? E9 de dema ku NRF2 tunebû hate dîtin. Ev bi nîşangirên otofagî yên guherandî re têkildar bû, di nav de rêjeyên fosfo-mTOR / mTOR û phospho-p70S6k / p70S6k (nîşana astengkirina otofagî), astên zêdekirî yên pêş-cathepsin D û hejmarek mezintir a laşên pirzimanî [119]. Di mişkan de APP-ya mirovî (V717I) û TAU (P301L) ku hev-îfade dikin, kêmasiya NRF2 bû sedema zêdebûna astên tevahî û phospho-TAU-yê di perçeya bêveng de û zêdebûnên APP-ên intraneuronal, digel hev kêmkirina astên neuronal ên p62, NDP52, ULK1, ATG5 û GABARAPL1. Co-herêmîbûn di navbera proteîna adapter p62 û APP an TAU de di nebûna NRF2 de kêm bû [36]. Bi tevahî, ev encam girîngiya NRF2-ê di otofagiya neuronî de radixin ber çavan.
Fakulteyên Karûbarên Celebên Têkiliyên Celeb Ji bo Hevrêziya Proteostasis
Di bin mercên dewletên dewlemend de, proteostasis bi rêya pêwendiyên protein-protein û guhertinên piştî-paşîn bi bersiva lezgîn ve tê kontrolkirin. Lêbelê, adaptasyona hucreyî hewce dike ku rêveberiya transcriptional ya jinên UPS, UPS û autophagy. Hişyariya hucreyên nervê bi tengahiyên zilmî yên kêm nizanin, bi tevlîheviya oksîdatîk û statotoxic, bi hêzkirina pêvajoya proteotasisê ji hêla modulasyonê veguherînal dibe dibe ku dibe ku alîkariya astengkirina mêjûyê bigirin.
Di rewşê de UPR, çalakiya her sê destan wê di encama veguhastina hin genes de (encam di [43] ve hate encam kirin. Ji bo nimûne, parçeyek ATF6-ê (ATF6f) bi bandorên bersivên ER-ê (ERSE) tête binivîse û behsa gelek genes, wekî XBPI, BIP û CHOP dihêle. Herweha, nîşana PERK-ê çalakiya faktoriya ATF4, dibe ku behsa behsa gelek cinsî ya UPR-ê û hinekên din jî di nav cinsên NRF2 de Hmox1 û p62 kontrol dikin. Di dawiyê de, encama çalakiya IRE1 di hilberîna faktorê veguherîn a çalak, XBP1 (XBP1s) de, ku kontrolkirina genes encoding protein di genya de protein de tevlî bibin.
Ji aliyê din ve, NRF1 ji bo mizgeftê ya gene ya proteînomal pêwîst e ku, di mêjûya Nrf1-knockout de diyar kir ku jenasên cûrbecî yên cûda yên 20S, û herweha tevlîhevkirina 19S bi tevliheviya proteasomal re bi xeletiyek kêmtir [90 ]. Hemî NRF1 û NRF2 ji bo celebên herêmên yên giyanên wê yên ku hene, pêşniyar dike ku çalakiyên pirtûkxaneyê rexne dike, lê çiqas ew bi awayekî di mekanîzmayên rêbaz û riya herêmî de [120] hene.
Fakslêdanên veguherînê yên bloka O ya (FOXO) malbata kontrolkirina genên xweser ên xweser ên xweser ên pirzimanî. Di heman demê de çi ye ku NRF2 bi pêvajoyê dibe, gelek layersên ku ji bo çalakiya FOXO, dibe ku li ser tedbîrên oksîterî yên xweserî anîn [121] hene. Di dawiyê de, faktora fonksiyonê TFEB, rêbazê masterê wekî biogenesis bi lîososomal tê fikra, di bin şertên tundûtûjiyê de xweseriya xweseriya xweseriya autophagy girîng e. Ji ber vê yekê, pêşangeha mTORC1 dibe ku ji bo TBBê û TFbedê ve veguherandina navberên xweser ên autophagy [122].
Bi tevahî, hebûna rêbazên veguherînan ên cihê yên van makîneran jî crosstalk û mekanîzmayên ku di bin rewşên cûda de cûreyên proteotasîsê bawer dikin dikin. Li gorî vê yekê, NRF2 dikare di nav destên girîng de têkildariyek girîng in ku piştgiriya asta oxidative bilind dibe. Ji bo nimûne, NRF2-ê zordestî-yê ku di bin şertên birçîbûnê de ji bo TFEB ji bo veguhastina veguhastina xweseriya dewlemend-ya xweser dike. Ji bilî, mizgefta mêjî di bin mercên dewlemend ên dewlemendî de, NRF2 wekî mekanîzmayek pêwendîdar e ku karûbarên autofagy di neuronan de çalak bike.
Ji bo NRF2 di Proteinopathiyan de Potansiyela Dermankirinê ya Hêzdar
Di çend salên dawî de pêşveçûneke mezin ya li ser rola rêveberiya UPR, UPS û autophagy li ser çalakiya NRF2, û hem jî ji bo veguhertina NRF2-yên pêkhênên van sê pergalên hevpeymaniyê. Ji ber vê yekê dibe, derfetên dermanên nû dikarin dibe ku ji bo betalkirina NRF2 ve di rêbazek girîng a protein de li ser nexweşî neurodegenerative.
Lêbelê, pirsek mayî ya sereke ev e ku ew ê kêrhatî an zirardar be ku hûn di mejî de asta NRF2 zêde bikin. Dibe ku analîzkirina daneyên epîdemolojîkî bersivek qismî bide, ji ber ku ew diyar dike ku gena NFE2L2 pir polorfîk e û hin polîmorfîzmayên nukleotîdê yên yekbûyî yên ku li herêma xweya birêkûpêkkerê de hatine dîtin dibe ku di asta nifûsê de û hin haplotipan de cûrbecûrbûna fîziolojîk di vegotina genê de peyda bike bi rîska kêmkirî û / an destpêkirina dereng a AD, PD an ALS re têkildar bûn [123]. Wekî din, wekî ku Hayes û hevalên wî nîqaş kirin [124], dibe ku bandora NRF2 bersivek U-teşe hebe, wateya ku astên NRF2 pir kêm dibe ku bibe sedema windabûna sîtoproteksyonê û zêdebûna hestiyariya stresker, dema ku pir NRF2 dikare hevsengiya homeostatîk ber bi senaryoyek kêmker (stresa kêmker), ku dê çewtî û berhevkirina proteînan favorî bike. Astên kêm NRF2 di mêjî de piştgirî dide ramanê ku hûrgelek hûrgelan dikare têr bibe ku di bin mercên patholojîk de sûd werbigire. Bi rastî, rola parastinê ya çalakkirina dermankolojîk a NRF2-navbeynkariya paqijiya proteînê di çanda hucreya neurodegenerasyonê ya cihêreng û di modelên vivo de hate xuyang kirin.
SFN çalakvanek dermankolojîk a NRF2 e ku hate xuyang kirin da ku îfadekirina genê proteasomal û xweseriyê pêk bîne [95], [36]. Balkêş e, Jo û hevalên wî destnîşan kirin ku SFN asta TAU-a fosforîlasyon kêm kir û Beclin-1 û LC3-II zêde kir, pêşniyar dike ku çalakkirina NRF2 dikare hilweşîna vê proteîna jehrîn bi riya otofagiyê hêsan bike [113]. Wekî din, hilweşîna mHtt bi SFN-ê hate zêdekirin, û ev bi karanîna MG132-ê hate vegerandin, nîşana hilweşîna proteasomal a vê proteîna jehrîn [95]. Têkbirina xweser a phospho- û ya bêserûber a bi autofagî re bi flavonoid organîk fisetin re hate ragihandin. Vê têkelê karibû autofagiyê biafirîne bi hevdemî çalakkirina û veguheztina nukleerî ya her du TFEB û NRF2, digel hin genên xweyên armanc. Ev bersiv ji hêla bêdengkirina TFEB an NRF2 ve hate asteng kirin [125]. Bott û hevalên wî ragihandin ku bandorên sûdwerbarê çalakvanek hemdem NRF2, NRF1 û HSF1 li ser jehrîbûna proteînê di atrofiya masûlkeyê ya spinal û bulbar de, nexweşiyek neurodegenerativ ku ji ber firehbûna CAG-a ku polîglûtamîn-kodkirin dubare dike ku tê de komikên proteîn hene [126]. Potansiyela aktîfkirina NRF2 ya ji bo dermankirina nexweşiyên neurodegenerativ bi pejirandina BG-12, formûlasyona devkî ya NRF2 dimetil fumarate (DMF), ji bo dermankirina skleroza multiple [127], [128] hate diyar kirin. Serkeftina DMF bi nexweşiyên otoîmmûn re ku bi pêkhateyek iltîhaba bihêz heye destnîşan dike ku nexweşiyên neurodegeneratif dikare ji veguheztina vê dermanê sûd werbigire. Di lêkolînek pêşîn klînîkî ya modela PD-ê de? -Sînukleînopatî de, hate xuya kirin ku DMF ji ber ku beşek e, ji ber pêgirtiya wê ya otofagî, neuroprotektîf e [129]. Lêkolînên ku bandorên sûdwergirtî yên NRF2 li ser neurodegenerasyonê radigihînin lê li ser bandora wê ya li ser paqijiya proteîn nesekinin hîn pirtir in (ji bo venêrînek berfireh, binihêrin [7]). Ev pir têkildar e, ji ber ku ew pir pêvajoyên zirarê radixe ber çavan ku dikarin bi hevdemî ji hêla yek lêdana NRF2 ve werin hedef girtin, di heman demê de stresa oksîdativ, neuroinflammasyon an fonksiyona mitokondrî jî tê de ye. Lêbelê, karê pêşerojê dê hewce be ku teqez were destnîşankirin ka aktîvkirina dermanolojîkî ya NRF2 dibe ku stratejiyek derbasdar be ku hilweşandina proteînên jehrî di mejî de hêsan bike.
Wekî ku berê jî hate şirove kirin, GSK-3 girantir kir? çalakî di nexweşiyên neurodegenerative de hate rapor kirin û ew tê texmîn kirin ku di encamê de kêmkirina NRF2 dikare bi qismî berpirsiyar be ji encama xirabkar re. Di bin van mercên patholojîk de, astengkerên GSK-3 dikarin hevkariyê jî bikin da ku astên NRF2 û proteostasis zêde bikin. Bandorên kêrhatî yên ragihankerên GSK-3 di modelên cihêreng ên nerodejenerasyonê de hatine rapor kirin û, balkêştir, zordestiya GSK-3 hate nîşandan ku asta proteînên jehrîn kêm dike [130], [131], [132], [133]. Her çend di navbera astengkirina GSK-3 û rêziknameya NRF2-transkrîpsiyonê ya genên ku proteostasis pêşve dibin de têkiliyên rasterast nehatine dîtin, lê maqûl e ku meriv texmîn bike ku daketina çalakiya GSK-3 dê bibe sedema zêdekirina astên NRF2, ku di dawiyê de dê bibe sedema bihêzbûnê proteostasis.
Çalakiya transkrîpsiyonê ya NRF2 û her weha kapasîteya hucreyî ya parastina proteostasis bi temenê re, faktorê xetereya sereke ya ji bo pêşvexistina nexweşiyên neurodegeneratif, kêm dibe. Ew maqûl e ku meriv bifikire ku xurtkirina NRF2 û, di encamê de, proteostasis dê, bi kêmanî, berhevkirina berhevokên proteîn û nerjenezerandinê dereng bixe. Birastî, dermankirina fibroblastên temenê mirovî bi 18? -Glycyrrhetinic acid (18? -GA) triterpenoid çalakkirina NRF2 pêş xist, dibe sedema derxistina proteasome û dirêjkirina temenê jiyanê. Vê lêkolînê destnîşan dike ku aktîfkirina dermankolojîk a NRF2 jî di emrê derengî de gengaz e [86]. Wekî din, lêkolînek paşê nîşan da ku ev terkîb li C.elegans bi navgîniya SKN-1 û çalakkirina proteasome bi bandorên bikêr li ser pêşveçûna AD-ê di modelên têkildar ên nematodê de [134] navbeynkarî kir.
Her tişt, fikrên NRF2-veguhestin yên genes yên proteotasis-related seemingly di proteinopathies de gelek fînanser e.
Sulforaphane û Bandorên Wê Li Ser Penceşêr, Mirin, Pîrbûn, Mêjî û Reftarî, Nexweşiya Dil & Zêde
Isothiocyanates hin hinek komên plankirî yên ku hûn di xwarinê de ne. Di vê vîdyoyê de ji bo wan ku ew her dem hatiye çêkirin pir girîng e. Hûrgelan kêmtir? Ji mijara yek ji xalên jêrîn bitikîne bi mijara xwe ya bijartinê vekin. Tiştek temamî bin.
Sernavên sereke
00: 01: 14 - Cancer û mirinê
00: 19: 04 - Aging
00: 26: 30 - Brain and behavior
00: 38: 06 - Reqameya dawî
00: 40: 27 - Dose
Full timeline:
00: 00: 34 - Destpêkkirina sulforaphane, fikirîn sereke ya vîdyoyê.
00: 01: 14 - Gelek mirin û kêmkirina xwarinê hemî mirinê.
00: 02: 12 - rîska kanserê Prostate.
00: 02: 23 - xetereya kansera kanserê.
00: 02: 34 - kansera lûksanê di kişandina xeteran de.
00: 02: 48 - Riskkirina kansera pêsîrê.
00: 03: 13 - Hypothetical: Heke ku hûn berê kanser heye? (interventional)
00: 03: 35 - mekanîzmayên mûzîzalîzmê daneyên danûstendinê yên kanser û mirinê.
00: 04: 38 - Sulforaphane û kanserê.
00: 05: 32 - belgeyên heywanan nîşan bide bandorkirina broccoli nîşan dide ku li ser rîskên tîrêjê di çilê de.
00: 06: 06 - Bandorkirina dermankirinê ya sulforaphane di nexweşxaneyên prostatê de.
00: 07: 09 - Biofilîzasyonê ya metotolyona di tissueya rastîn de.
00: 08: 32 - Pêşangeha kanserên stenbolê stem cells.
00: 08: 53 - Dersa Dîroka: Brassicas wekî xaniyên tendurustî jî di Romayê kevnar de ava kirin.
00: 24: 14 - Xwendekarên Mouse-ê pêşniyar dike ku sulforaphane dibe ku karibe parastinê ya di temenê temenê de çêtirîn.
00: 25: 18 - Sulforaphane pêşveçûna porê çêtirîn di modela mîkrobê de balê. Wêne li 00: 26: 10.
00: 26: 30 - Destpêkkirina mêjû û tevgerên sektorê.
00: 27: 18 - Bandora tesîrê ya broccoli derkeve ser autism.
00: 27: 48 - Bandora glucoraphanîn li schizophrenia.
00: 28: 17 - Destpêkiya dîskêşiyê (mekanîzmayên mektezîzmê û xwendevanan).
00: 31: 21 - Xwendina Mûxoyê bi karanîna xnumxên cuda cuda cuda yên tundûtûjî-stêrdekirin nîşan dide ku sulforaphane bi awayekî fluxoxetine (prozac).
00: 32: 00 - Lêkolînek rasterast ya glucoraphanîn lêkolînek di heman demê de di heman demê de di heman demê de deverek bi bandorkirina astengiya ji modela stratejiya sosyal ve.
00: 33: 01 - Destpêkirina beşa neurodegenerasyon.
00: 33: 30 - Nexweşiya Sulforaphane û Alzheimer.
00: 33: 44 - Nexweşiya Sulforaphane û Parkinson.
00: 33: 51 - Sulforaphane û nexweşiya Hungtington.
00: 35: 01 - Sulforaphane lezgîn piştî ku TBI ji bo bîranîn (xwendina mûsa) çêtir dike.
00: 35: 55 - Sulforaphane û plastîkaya neuronal.
00: 36: 32 - Sulforaphane hînbûna modela şertê duyemîn diabetesê de çêtir dike.
00: 37: 19 - Sulforaphane û duchenne dystrophy.
00: 37: 44 - Myostatin li pêşangehên hucreyên satelê (li vitro).
00: 38: 06 - Daxistina nerazîbûnê û kanserê, zirarê DNA, zexta oksîtatiyê û tehlîzanê, benzene xemgîniyê, nexweşiya cardiovascular, cureyê diyabetê, bandorê li ser mêjî (bandor, xweserîzmê, schizophrenia, neurodegeneration), riya NRF2.
00: 40: 27 - Fikir li ser doktorên broccoli an sulforaphane.
00: 41: 01 - Li ser malbata dorpêçkirin.
00: 43: 14 - Li ser karkirina xwarinê û çalakiya sulforaphane.
00: 43: 45 - Bacteriya guhertinê ya sulforaphane ji glucoraphanin.
00: 44: 24 - karê karên çêtir çêtir dibe gava ku bi tevgera myrosinase ve ji aliyên veberhênanê re hev.
00: 44: 56 - teknolojiyên xwarinê û sebên zêrîn ên xerîb.
00: 46: 06 - Asotiocyanates wek goitrogens.
Faktoriya nukleer a erythroid-derived 2 (NF-E2) -iya 2-ê, nehêle wekî Nrf2 tê naskirin, faktoriyek veguherînek e ku behsa xweseriya antioxidant û binavkirina enzyme dike. Lêkolînên Lêkolînan jî di bin kontrola oxuxative kontrola xwe de nîşan dide. Piraniya nexweşiyên neurodegenerative, wekî nexweşiya Alzheimer û nexweşiya Parkinson, bi xemgîniya oksîdative û sermayona kevneşopî, armanca hevpar ên Nîqaşên nrf2. Dr. Alex Jimenez DC, CCST Insight
Şîrovekirinên dawî
Faktora veguhastinê NRF2 anchestrates bersivên pêvajoya proteostatic bi veguherandina guhertin û guhertinên di UPR, UPS û autophagy (Fig. 4). Ji ber vê yekê, nebûna kêmbûna NRF2 hatiye nîşandan ku proteinopathy aggravate, pêşniyar dike ku NRF2 ji bo paqijiya proteîn a herî girîng e. Bi hev re, em dikarin bifikirin ku NRF2 ji bo proteinopathies armancên dermankek balkêş be.
4 NRF2 Nîşanek wek hub-nîşanên pêvajoyên prototoxic-derveyî ji bo bersivê veguhastina parastina rewa ve girêdayî ye. Gelek pêkanîna nebaş / proteînên nefolded dê di çalakiya ER-ê de bersivê hilberîna protein-aşkere (UPR) dibe. Çalakiya PERK an MAPK dibe ku di encama veguherîna transcriptal de Gpx8-ê û gelek enzyme dibe ku GSH-asta veguherînin, ji bo pêkanîna protein-çermê rast bikin. Komên proteînan çalakiya proteasome (UPS) derxistin, dibe ku ji hûrgelên NRF2 vekin. NRF2 hatiye nîşandan ku bi taybetî veguherîna pelma Psma3, Psma6, Psmb1, Psmb5 û Pênesên Pomp. Gelek dabeşeyên din ên di bersiva NRF2-ê de bi bersiva D3T ve hatine birêvekirin, dibe ku ji lîsteya NRF2 de lîsteya hilberên proteasokên mezintir dibe. Autophagy ji bo hilweşîna proteîneyên kevneşopên rêberê sereke ye. Autophagy jî NRF2 veguherîne, bi rêvekirina vê dezgeha NRF2 bi p62, Ndp52, Ulk1, Atg2b, Atg4c, Atg5, Atg7 û Gabarapl1.
Li gor gotara jorîn, dema ku nîşanên nesaxên neurodegenerative bi riya alternatîfên cûre derman kirin, lêkolînên lêkolînan nîşan didin ku çalakiya Nrf2 dikare nêzîkbûnek tedawî be. Bo Çalakvanên Nrf2 bi armancên mekanîzmayên mezin ên nexweşî hedef bikin, hemî nexweşiyên nerjeneger dikarin ji karanîna faktora transkrîpsiyonê Nrf2 sûd werbigirin. Dîtinên Nrf2 şoreşa dermankirina nexweşiyên neurodegenerative veguherandiye. Berfirehiya agahdariya me bi pirsgirêkên tenduristiya chiropractic û spinal ve girêdayî ye. Ji bo nîqaşkirina mijarê, ji kerema xwe ji Dr. Jimenez bipirsin an jî bi me re têkilî bikin at 915-850-0900.
Pirsgirêk Pirsgirêk: Pevçûnê Gelek Kuştina bêyî Surgery
Painşa çokan semptomek navdar e ku dikare ji ber cûrbecûr birîndarbûn û / an şert û mercên çokan çêbibe, tê de birînên werziş. Çok di laşê mirov de yek ji hevgirêkên herî tevlihev e ji ber ku ji hevberdana çar hestî, çar lîgament, tendonên cihêreng, du menisci û kartilan pêk tê. Li gorî Akademiya Amerîkî ya Bijîjkên Malbatê, sedemên herî gelemperî yên êşa çokê di nav de binavbûna patellar, tendinitis patellar an çokê jumper, û nexweşiya Osgood-Schlatter hene. Her çend êşa çokê bi îhtîmalek mezin di mirovên ji 60 salî mezintir de çêbibe jî, êşa çok dikare di zarok û ciwanan de jî çêbibe. Êşa çokê li gorî rêbazên RICE dikare li malê were derman kirin, lêbelê, birînên giran ên çokê dibe ku guhdana bijîjkî ya tavilê hewce bike, tevî lênihêrîna chiropractîk. �
Zextên oksîterî tête ku wekî zirarê hucreyê ji hêla radîkayên serbixwe ve, an jî ankek neserable, ji dawiyê bi karê tendurustî bandor dike. Cenazeyê mirovî radikên serbixwe dike ku ji bacteria û viruses neutralize nebe, lêbelê, faktorên derveyî, wekî oxygen, qezenc û radiasyonê, pir caran dikarin radîkal azad bikin. Stenbolê ya oksîterî gelek pirsgirêkên tenduristiyê ve girêdayî ye.
Stress û pisporên din ên zordestî li ser parastina mekanîzmayên navxweyî veguherînin ku dikare alîkariya bersiviya antioxîdant a laşê saz bikin. Nrf2 proteîn e ku asta asta oxaxative stressê dike û hucreyên ku ji hêla faktorên hundur û hundir biparêze bikin. Nrf2 jî ji bo ku di hilberîna antîxîdantên enzymes û zextên rehber-bersivê de beşdarî tevlî cinsên zagonî birêve bike. Armanca jêrîn jêrê ye ku ji bo şirovekirinê ye bandorên Nrf2 di kansera
Abstract
Rêberê Keap1-Nrf2 rêveberê sereke ya bersivên cytoprotîk e ku zextên oksîdatîf û elektrofîlîk e. Tevî ku rêyên rêyên rêyên rêbazên ku ji hêla faktora veguhestina Nrf2 ve di destpêka pêşveçûn û pêşveçûnê de di bin destûra normal û pêşerojên paşîn de, di hucreyên temamî de nrf2 çalakiya pêşveçûnê ya karmendê û kansera tûlor zêde dibe. Di vê pevçûnê grafîk de, em li ser hucreyên kansera kanseriyê yên Keap1-Nrf2 û dîsregulasyona wê pêşkêş dikin. Em jî kurtasî encamên encamên nucf2 di asta kanserên kansera kanser de û kurteya tedawiya gene di kanserê de bikar bînin.
Keywords:Nrf2, Keap1, Cancer, Alîkariya Antioxidant, Gene Therapy
Pêşkêş
Riya Keap1-Nrf2 sererastkerê sereke yê bersivên sîtoprotektîf ê li stresên endojen û dervayî yên ji hêla cinsên oksîjena reaksiyonî (ROS) û elektrofîl ve têne çêkirin e [1]. Proteînên girîng ên nîşankirinê di rê de faktorê veguhastinê Nrf2 (faktorê nukleerî yê têkildar erythroid 2-pêwendîdar 2) e ku bi proteînên Mafê piçûk re bi hev re bi hêmana bersiva antioxidant (ARE) ve girêdayî ye li herêmên birêkûpêk ên genên armanc, û Keap1 (Kelch ECH girêdana proteîna 1), proteînek zordar ku bi Nrf2 ve girêdide û bi rêça ubiquitin proteasome hilweşîna wê pêşve dibe (Hêjîrê. 1). Keap1 proteînek pir dewlemend a cysteine e, Keap1 mişk bi tevahî 25 û 27 mirov bermahiyên cysteine hene, ku piraniya wan dikare di vitro de ji hêla oksîdan û elektrofîlên cihêreng ve were guhertin [2]. Sê ji van bermahiyan, C151, C273 û C288, hate xuyang kirin ku bi guherîna lihevhatina Keap1 ku dibe sedema veguheztina nukleerî ya Nrf2 û derbirîna genê hedefê ya paşîn [3] (Hêjîrê 1) rolek fonksiyonel dilîzin. Mekanîzmaya rastîn a ku guhertinên cysteine-ê di Keap1-ê de dibe sedema çalakkirina Nrf2-ê nayê zanîn, lê du modelên serdest lê ne bi veqetandî yên hevûdu ne (1) modela hinge û latch, ne, ku tê de Guherandinên Keap1 di bermahiyên tiyolê de yên di IVR ya Keap1 de dimînin têkiliya bi Nrf2 re têk bibe sedema nerastkirina bermahiyên lîzîn di nav Nrf2 de ku nema dikare were polyubiquitinylated û (2) modela ku tê de guhertina tiyol dibe sedema veqetandina Cul3 ji Keap1 [3]. Di her du modelan de, Keap2 ya ku bi nêrgizê ve hatî guhertin û bi Nrf1 ve girêdayî ye neçalak e û, di encam de, proteînên Nrf2 yên nû sentezkirî Keap1-ê dorpêç dikin û dikevin nav navikê, bi ARE-yê ve girêdidin û vegotina genên armancên Nrf2-ê wekî NAD (P) H diavêjin quinone oxidoreductase 1 (NQO1), heme oksîjenaza 1 (HMOX1), glutamate-cysteine ligase (GCL) û glutathione S transferazên (GST) (Hêjîrê 2). Ji bilî guheztinên tiyolên Keap1 ku di encamê de Nrf2 gene hedef digire, proteînên wekî p21 û p62 dikarin bi Nrf2 an Keap1 ve girêbidin bi vî rengî têkiliya navbera Nrf2 û Keap1 têk bibin [1], [3] (Hêjîrê. 3).
Hêjmar 1. Avahiyên Nrf2 û Keap1 û koda cysteine. (A) Nrf2 ji 589 asîdên amînoyî pêk tê û şeş warên evolusyonî yên pir hatine parastin, Neh1-6. Neh1 motîfek bZip, deverek bingehîn a structure leucine zipper (L-Zip) heye, ku devera bingehîn ji naskirina DNA berpirsiyar e û L-Zip bi proteînên Mafê yên piçûk dimerîzasyonê dike. Neh6 wekî degron kar dike ku di navgîniyê de hilweşîna Nrf2 navbeynkariyê bike. Neh4 û 5 warên veguhastinê ne. Neh2 motîfên ETGE û DLG hene, ku ji bo têkiliya bi Keap1 re, û herêmek hîdrofîlî ya bermahiyên lîzîn (7 K), ku ji bo polîbibijandin û hilweşîna Nrf1 a girêdayî Keap2 pêdivî ne, pêdivî ye. (B) Keap1 ji 624 bermahiyên amîno asîd pêk tê û pênc warên wê hene. Du motîfên têkiliya proteîn-proteîn, qada BTB û qada Kelch, ji hêla herêma navbeynkar (IVR) ve têne veqetandin. Domana BTB digel beşa N-termînalê ya IVR navbeynkariyê dike homodimerîzasyona Keap1 û bi Cullin3 (Cul3) ve girê dide. Domana Kelch û herêma C-termînalê bi Neh2 re têkilî danî. (C) Nrf2 bi motîfên Neh1 ETGE û DLG bi du molekulên Keap2 re têkiliyê datîne. Herdu ETGE û DLG li ser rûyê jêrîn ê motifa Keap1 Kelch bi malperên wekhev ve girêdidin. (D) Keap1 di bermahiyên cysteine de, bi 27 cysteine di proteîna mirovan de dewlemend e. Hin ji van cysteines li nêzê bermayiyên bingehîn cih digirin û ji ber vê yekê hedefên hêja yên elektrofîl û oksîdan in. Nimûneya guherînê ya bermayiyên cysteine ji hêla elektrofîl ve wekî koda cysteine tê zanîn. Hîpoteza koda cysteine pêşniyar dike ku ajanên aktîfker ên Nrf2 bi avakî cûda bandorê li cîsteînên Keap1 ên cuda dikin. Guherandinên cysteine dibe sedema guhertinên lihevhatî yên Keap1 ku têkiliya di navbera warên Nrf2 DLG û Keap1 Kelch de têk dide, bi vî rengî polyubiquitination-a Nrf2 asteng dike. Girîngiya karbidest a Cys151, Cys273 û Cys288 hate xuyang kirin, ji ber ku Cys273 û Cys288 ji bo tepisandina Nrf2 û Cys151 ji bo aktîfkirina Nrf2 ji hêla induktoran ve hewce ne [1], [3].
Nig. 2. Nrf2-Keap1 signaling pathway. (A û B) di rewşên basal de, du du keap1 molekulên girêdayî Nrf2 û Nrf2 ji alîyê Cul3-based E3-ê veguhestî ye. Ev encamên polîtîkûyîtîk di rapidê Nrf2 de hilweşandina hilberîna proteasome. Nêzikek biçûk ya Nrf2 ji berhevkirina kozîkan ve direve dike û di nav nucleus de berbi navnîşî ya navendî ya navxweyî ya girêdayî jene girêdayî ye, bi vî awayî hûrgelê malostasiyan biparêzin. (C) Di bin mercên zordariyê de, hilberên guhdarên Keap1 cysteines berfireh bikin ku bi pêşniyariya Nrf2-ê veguhestina dorpêçê ya tevlîhevî. (D) Li gor nimûne û pişkek laş, guhertina taybetmendiyên Keap1 cysteine di rêgezên nû de li Keap1 encam kirina encamkirina Nrf2 DLG ji Keap1. Berbiçavkirina Nrf2 tête kirin lê belê bi bendava bi motora ETGE dimîne. (E) Di modelê de Keap1-Cul3 dabeşkirina hûrgelan, bendkirina Keap1 û Cul3 di bersivên elektrofîlan de, ji ber ku ji rêberê Nrf2 ve ji pergala mubiquitandinê ve tê veşartin. Di modela pêşniyarên pêşniyazên pêşniyaz de, aktor-guhertin û Nrf2-bound Keap1 ne diqewimin e û di encamê de, di encama nû de, peymanên nû Nrf2 bi rêberên Nrf1 Keap2 û nucleus veguherîne, li hember Nucleya Antioxidant Element (BERSÎVE) û behsa derbazkirina Nrf1 veguherîne genên wek NQO1, HMOX1, GCL û GSTs [3], [XNUMX].
Nig. 3. Mîkrozmayên ji bo kombûna navokî ya Nrf2 di kanserê de. (A) Mutavên Somatîk ên di Nrf2 an Keap1 de têkiliya van du proteins. Di Nrf2 de, mutasyonên bandorên ETGE û DLG bandor dike, lê di navên Keap1 de bêtir belav kirin. Ji bilî, çalakiya îkogogene, wekî KrasG12D [5], an astengên tromor ên astengî, wekî PTEN [11] dibe ku veguherandina veguherîna Nrf2 û di nav Nrf2 de zêde dibe. (B) Hypermethîlasyonê ku protoktorê Keap1 di lung û kanserê prostatê de dibe sedema kêmkirina Keap1-MRNA, ku ji berhevkirina navokî ya Nrf2 zêde dibe [6], [7]. (C) Di parsiyonê ya malbata gerîla carcinoma, wenda dibe ku çalakiya enzyme hydrajîtê ya fumarateyê ji berhevkirina fumaratê û bêtir ji bo teslîmkirina Keap1 Cysteine residues (2SC) dibe. Guhertina vê post-wergerê ji bo astengkirina Keap1-Nrf2 veguhastin û kombûna navokî ya Nrf2 [8], [9]. (D) Bihevkirina hilberên protokolên wek p62 û p21 dikare Nrf2-Keap1 tengahî û encam dibe ku di zêdebûna Nrf2 ya nukleerî de dibe. p62 bi keap1 veguhastina pocket ji bo Nrf2 û p21 bi rasterast bi rasterast bi têkiliyên DLG û ETGE yên Nrf2 ve girêdayî ye, bi vî awayî Keap1 re [10].
Mîkroozîsyona Çalakî û Dysregulation ya Nrf2 di Cancer
Tevî ku cytoprotection ji hêla çalakiya Nrf2 ve tête darizandin ji bo kanseriya normal û atîkalên paşîn, di hucreyan de temamî temamî girîng e girîng e ku çalakiya Nrf2 bi zêdebûna karmendiyê kansera kanseriyê û zêdebûna hucreyê tumorê [4] dide. Gelek mekanîzmayên ku bi rêberiya Nrf2 rêvegirtina rêwîtiyê ye, di kelepên cuda de têne çalak kirin, tête diyar kirin: (1) li ser Keap1 an Kap1 ya Rêveberiya Keap2 ya Nrf2 veguherîna wan veguherand; (1) şikandina epagenetîk ya Keap2 expressionên ku ji zordestiya nerast a Nrf3; (62) ji bo proteinên dîktatorî yên wekî p1 ku ji ber tevlihevkirina Keap2-Nrf4 complex; (2) veguherandina veguherandina Nrf5 ji hêla oncogenic K-Ras, B-Raf û c-Myc; (1), [3], [4], [5], [6], [7], [8], [9], [10], [3], [2], ji [xNUMX] veguherandina post-translationê ya post-translational ya keap2 cysteinesên ku di navberiya papalaya nermalavê de, 4], [2], [11] (Nig 4). Pêvek pêkhatî ya NrfXNUMX dibe sedema şirovekirina genên zêdebûna jînabolîzmê ya ku di vî awayî de berxwedana zêdebûna dermanên chemotherapeutîk û radyotrîkê. Herweha, asta proteîn ya NrfXNUMX bilind bi bi kansera nexweşî ya nexweşî ve girêdayî ye [XNUMX]. Pirrjimar NrfXNUMX jî bi bandorkirina hucreyê guluco û glutamine bi rêberiya rêbazên anabolic reklama rêkûpêk û pentose fosphate bandora bandorkirina hilberîna hucre [XNUMX] (XNUMX).
Nig. 4. Roja dual ya Nrf2 di tumorigenesis de. Di bin mercên fîzîkî de, asta nizanîkî Nrf2 ji bo parastina malê malperasis in. Nrf2 ji hêla carcinogens, ROS û ajokarên din ên DNA-yên din ên hilweşîna destpêkirina tumor destpêkirin û kansera metastasis ve dike. Di tumorigenesisê de, zirarê DNA-ê dihêle zordariyê ya Nrf2-ê ku hucreyên xweser ên xweser ên xweser ên ku alîkarî ROS-ê ya endojen ROS-ê dike û ji bo apoptosis dûr nekin. Asta nukleer a Nrf2 bi awayekî jînavî çalak bikin, tevî cenawirên cytoprotîk ên beşdarî xwarina hilberîner û metrokola hûrgelan zêde kir. Astengdarên ku bi bilindtirîn Nrf2 ve girêdayî nexweşî ne belengî ne ji ber ku ji radyoya kemerografîk û hilberîna kansera kanserê aggressive ve girêdayî ye. Bi vî awayî, çalakiya rêwîtiya Nrf2 di destpêkirina qonaxên tumorigenesis de parastî ye, lê di paşînan de paşê xera dikin. Ji ber vê yekê, ji bo parastina kanserê, Kurtkirina çalakiya Nrf2 veguhestineke girîng e, lê ji bo dermankirina kansera Nrf2, pêdivî ye. [4], [11].
Given ku çalakiya Nrf2 bi gelemperî di hucreyên kanseran de digel encamên nerazîbûnê pêk tê, hewce ne hewce ye ku tedawiyên Nrf2. Mixabin, ji ber ku wekheviya nermalavî digel hin kesên bZip ên malbatî, pêşveçûnên taybet ên Nrf2-ê yên balkêş e ku xebitînek dijwar e û tenê çend lêkolînan ji hêla Nrf2 ve hate belavkirin. Bi hilberên xwezayî yên firotanê, Ren et al. [12] wekî brusatol wekî nrf2 bêdengdar e ku pisporên kemotîktutîk ên cisplatin bilind dike. Ji bilî, PI3K nexşandin [11], [13] û Nrf2 siRNA [14] bikar anîna ku di hucreyên kanserê de Nrf2 veguherînin. Di demê dawiyê de, me nêzîkek alternatîf bikar anîn, wekî wekî tedawiya xwekujiya kansera kanseriyê tê zanîn, da ku hucreyên kansera bi bilindkirina Nrf2 bilind bikin. Nrf2-powered veguhên lentiviral [15] ku bi kîmeya thymidine (TK) ve tête nav hucreyên kansera bilind ên li RI-ê ve ye û hucreyan bi pro-derman, ganciclovir (GCV) têne derman kirin. GCV bi metabolîzasyona bi GCV-monophosphate ye, ku ji hêla fosphorylîteyê ve bi hêla kêlîkên hucreyê ve tête nav forma sêyemîn a tifphosphate a zext (16) (Fig 5). Ev yek dibe sedema kuştina bandorên bandor ên ne tenê TK bi hucreyên tîmor hene, lê herweha hucreyên cîran ên ji ber bandorê dorpêçê [17]. TK / TCV ya GCV-ê ji hêla dermankirina kanserê kaymotherapeutic doxorubicin ve tê dermankirin [16], piştevaniya piştevanîya ku ev nêzîk dibe ku bi rêbazên bi kevneşopî yên kevneşopî re bikêr bibin.
Nig. 5. Tedawiya xwekujî. Destûra Nrf2 di asta hucreyan de di nav hucreyên kanserê de dikarin bi karanîna Nrf2-veguherîna viral vector ji bo xwekêşana xwekujî ya xwenîşandanê [16]. Di vê helwestê de, veguhestina lentiviral (LV) di bin qeşeyên SV40-ê de bi kêmtirîn kîmîteya thymidine (TK) tê gotin ku bi çarçoveyên çar herêman bi hucreyên adenocarcinoma ve têne veguhestin. Rêjeyên nukleer ên Nrf2 bi rêberkirina axaftina TK ya bi Nrf2 re berbiçav dibe. Hingê hucreyên ku ji hêla TK-ê, ganciclovir (GCV) ve tê tedawî kirin, ji hêla TK-ê ve tê fosphorylated. GCV Têkiliya DNA veguherîne û dibe sedema kuştina bandor ê bandor ne tenê ne tenê TK bi hucreyên tîmor hene, lê hucreyên cîran jî ji ber ku bandora bandor.
Nrf2 ku rêveberiya masterê ye ku ji ber laşê mirovî ya ku di alîkariya tedawiya oksîterê de hilweşîne hilberîna hilberîna antîxîdîdantan dike. Enzymes yên gelek antîxîdant, wekî wekî dermanên superoxide, an SOD, glutathione û catalase, di rêya Nrf2 de jî diçin jî. Ji bilî, hin phytochemîkên mîna turmerîk, ashwagandha, bacopa, çayek kesk, û şîrê şîrê, Nrf2 çalak bikin. Lêkolînên lêkolînê hene ku Çalakiya Nrf2 dikare bi xwezayî dikare parastina sehêlê û zêdebûna laşê mirovî re bistînin.
Dr. Alex Jimenez DC, CCST Insight
Sulforaphane û Bandorên Wê Li Ser Penceşêr, Mirin, Pîrbûn, Mêjî û Reftarî, Nexweşiya Dil & Zêde
Isothiocyanates hin hinek komên plankirî yên ku hûn di xwarinê de ne. Di vê vîdyoyê de ji bo wan ku ew her dem hatiye çêkirin pir girîng e. Hûrgelan kêmtir? Ji mijara yek ji xalên jêrîn bitikîne bi mijara xwe ya bijartinê vekin. Tiştek temamî bin.
Sernavên sereke
00: 01: 14 - Cancer û mirinê
00: 19: 04 - Aging
00: 26: 30 - Brain and behavior
00: 38: 06 - Reqameya dawî
00: 40: 27 - Dose
Full timeline:
00: 00: 34 - Destpêkkirina sulforaphane, fikirîn sereke ya vîdyoyê.
00: 01: 14 - Gelek mirin û kêmkirina xwarinê hemî mirinê.
00: 02: 12 - rîska kanserê Prostate.
00: 02: 23 - xetereya kansera kanserê.
00: 02: 34 - kansera lûksanê di kişandina xeteran de.
00: 02: 48 - Riskkirina kansera pêsîrê.
00: 03: 13 - Hypothetical: Heke ku hûn berê kanser heye? (interventional)
00: 03: 35 - mekanîzmayên mûzîzalîzmê daneyên danûstendinê yên kanser û mirinê.
00: 04: 38 - Sulforaphane û kanserê.
00: 05: 32 - belgeyên heywanan nîşan bide bandorkirina broccoli nîşan dide ku li ser rîskên tîrêjê di çilê de.
00: 06: 06 - Bandorkirina dermankirinê ya sulforaphane di nexweşxaneyên prostatê de.
00: 07: 09 - Biofilîzasyonê ya metotolyona di tissueya rastîn de.
00: 08: 32 - Pêşangeha kanserên stenbolê stem cells.
00: 08: 53 - Dersa Dîroka: Brassicas wekî xaniyên tendurustî jî di Romayê kevnar de ava kirin.
00: 24: 14 - Xwendekarên Mouse-ê pêşniyar dike ku sulforaphane dibe ku karibe parastinê ya di temenê temenê de çêtirîn.
00: 25: 18 - Sulforaphane pêşveçûna porê çêtirîn di modela mîkrobê de balê. Wêne li 00: 26: 10.
00: 26: 30 - Destpêkkirina mêjû û tevgerên sektorê.
00: 27: 18 - Bandora tesîrê ya broccoli derkeve ser autism.
00: 27: 48 - Bandora glucoraphanîn li schizophrenia.
00: 28: 17 - Destpêkiya dîskêşiyê (mekanîzmayên mektezîzmê û xwendevanan).
00: 31: 21 - Xwendina Mûxoyê bi karanîna xnumxên cuda cuda cuda yên tundûtûjî-stêrdekirin nîşan dide ku sulforaphane bi awayekî fluxoxetine (prozac).
00: 32: 00 - Lêkolînek rasterast ya glucoraphanîn lêkolînek di heman demê de di heman demê de di heman demê de deverek bi bandorkirina astengiya ji modela stratejiya sosyal ve.
00: 33: 01 - Destpêkirina beşa neurodegenerasyon.
00: 33: 30 - Nexweşiya Sulforaphane û Alzheimer.
00: 33: 44 - Nexweşiya Sulforaphane û Parkinson.
00: 33: 51 - Sulforaphane û nexweşiya Hungtington.
00: 35: 01 - Sulforaphane lezgîn piştî ku TBI ji bo bîranîn (xwendina mûsa) çêtir dike.
00: 35: 55 - Sulforaphane û plastîkaya neuronal.
00: 36: 32 - Sulforaphane hînbûna modela şertê duyemîn diabetesê de çêtir dike.
00: 37: 19 - Sulforaphane û duchenne dystrophy.
00: 37: 44 - Myostatin li pêşangehên hucreyên satelê (li vitro).
00: 38: 06 - Daxistina nerazîbûnê û kanserê, zirarê DNA, zexta oksîtatiyê û tehlîzanê, benzene xemgîniyê, nexweşiya cardiovascular, cureyê diyabetê, bandorê li ser mêjî (bandor, xweserîzmê, schizophrenia, neurodegeneration), riya NRF2.
00: 40: 27 - Fikir li ser doktorên broccoli an sulforaphane.
00: 41: 01 - Li ser malbata dorpêçkirin.
00: 43: 14 - Li ser karkirina xwarinê û çalakiya sulforaphane.
00: 43: 45 - Bacteriya guhertinê ya sulforaphane ji glucoraphanin.
00: 44: 24 - karê karên çêtir çêtir dibe gava ku bi tevgera myrosinase ve ji aliyên veberhênanê re hev.
00: 44: 56 - teknolojiyên xwarinê û sebên zêrîn ên xerîb.
00: 46: 06 - Asotiocyanates wek goitrogens.
Spasî
Ev xebata bi Akademiya Fînlandê, Weqfa Sigrid Juselius û Rêxistinên Kanser ên Fînîstî hate piştgir kirin.
Di encamê de, faktora nukleerê (erythroid-2-werdigire 2) -wek 2, wekî NFE2L2 an Nrf2 jî tête zanîn, proteînek e ku hilberîna antioksîdan zêde dike ku laşê mirov li hember stresa oksîdatîf diparêze. Wekî ku li jor hate vegotin, teşwîqkirina riya NrfXNUMX ji bo dermankirina nexweşiyên ji ber stresa oksîdativ, di nav de penceşêr de jî têne xwendin. Berfirehiya agahdariya me bi pirsgirêkên tenduristiya chiropractic û spinal ve sînor e. Ji bo nîqaşkirina mijarê, ji kerema xwe ji Dr. Jimenez bipirsin an jî bi me re têkilî bikin at 915-850-0900.
Pirsgirêk Pirsgirêk: Pevçûnê Gelek Kuştina bêyî Surgery
Painşa çokan semptomek navdar e ku dikare ji ber cûrbecûr birîndarbûn û / an şert û mercên çokan çêbibe, tê de birînên werziş. Gelek yek ji nav laşên mirovahiyê ye ku ev yek ji hestê çar çar hestî, çar ligaments, celebên cûda, du mêran û kartilan e. Li gorî Akademiya Xweseriya Danîmarkî ya Amerîkayê, sedemên gelemperî yên tehlûkê yên herî tehlûkê di nav deverên patellar, pirtûka pirtûştê an jî gumperê jumper, û nexweşiya Osgood-Schlatter hene. Tevî ku tehdîtê çermê pir zêde dibe ku li ser 60 salî di nav zarok û ciwanan de dibe ku dibe ku gefê xwînê. Pêwîstiya RICE di malê de piştî rêbazên RICE tê dermankirin, lêbelê, birîndarên giran ên giran dikarin hewceyê baldarî dermanî, bi lênêrîna chiropractîk re.
DNA bi hema 20,000 genes piştgirî dike, her bernameyeke bernameyek ji bo proteînek an enzyme hewce dike ku ji bo jiyanek tendurustî hewce dike. Her yek ji van pîvanan hewce ne ku bi awayekî "promoter" têne birêvebirin, ku bi rastî tête ku çiqas kîjan materyal û / an kîmyewî tête kirin û di bin şertên wê de jî pêşve bibin.
Bi vekirina girêdana taybetî ya rengê wekheviya pêşveçûnê yên wekî promoter, ku wekî wekî Antioxîdant Response Element, tê zanîn, an Nrf2 factorLeza afirandinê ji bo bi sedan genên cihêreng piştgirî dike ku dihêle şaneyan di bin şert û mercên stresê de bijîn. Dûv re van genan hilbijarkek enzîmên antîoksîdan çêdikin ku bi bêbandorkirina oksîdantan û bi paqijkirina hilberên jehrîn ên ku di hilberîna wan de mane, torgilokek parastinê pêşve dixin, ji bilî ku alîkariya vegerandina zirara ku wan kirine.
Astengiya Oxsyayî çi ye?
Gelek oksîdanên mîna radîkalê superoksîdê, an O2-., û hîdrojen peroksîtê, an H2O2, bi pratîka şewitandina maddeyên û/an kîmyewî yên ku laşê mirovan diparêzin hatine afirandin. Laşê mirov xwedan enzîmên antîoksîdan e ku xwarin û vexwarinên reaktîf ên ku em dixwin bêbandor dikin û ji holê radikin. Nrf2 hilberîna wan modul dike da ku hevsengiyê biparêze û daxwaziya van hemî enzîman destnîşan dike. Ev hevsengî dikare ji hêla çend faktoran ve, di nav de temen, were qut kirin.
Her ku em kal dibin, laşê mirov kêmtir Nrf2 diafirîne û ev hevsengiya nazik dikare hêdî hêdî dest pê bike ku ber bi aliyê oksîtasyonê ve bizivire, rewşek ku wekî stresa oksîdative tê binav kirin. Nexweşî jî dibe sedema hilberîna zêde ya oksîdantan. Enfeksiyon, alerjî, û nexweşiyên otoîmmune jî dikarin hucreyên meya xweparastinê biafirînin ku oksîdanên reaktîf, wek O2- biafirînin. , H2O2, OH û HOCl, ku hucreyên saxlem xera dibin û bi iltîhaba bersiv didin. Nexweşiyên ku bi pîrbûnê re têkildar in, di nav de êrîşên dil, stok, kansera, û şertên neurodejenerative yên mîna nexweşiya Alzheimer, di heman demê de pêşveçûna oksîdantan jî zêde dikin, stresê û bersivek iltîhaba çêdikin.
Nrf2 Çalakvanan çi ne?
Pro protein ya Nrf2, ji hêla ku enzymes û genes dikare piştevanî û kontrolkirina rêbazê veguherandiye, ji hêla reaksiyonê biochemîkî ve di nav hucreyê de ku di guherîna guhertinê de di nav cûdaweriya giyanî û herweha wekheviya oksîtîvî ye. Hûrgelên hestiyar ên vê rêwrê biguherînin û Nrf2 veguherîne, dibe ku bi vî awayî dibe ku ew di nav hucreyê ya hucreya DNA de belav bibe. Nrf2 dibe ku alternatîf bi veguherîne an genes û enzîmên veguherînin ku ji bo parastina hucreyê piştgirî dike.
Bi kêfxweşî, cûrbecûrên cûrbecî ku çalakvanên Nrf2 di rêya xwarina hin hûrgelan de têne çêkirin û pêşveçûna sedsala berê de di peymana kevneşopî ya kevneşopî yên Çînî û Niştimanî ya karanîna karanîna karanîn. Ev phytochemîkan wekhevî wekî bi kêmtir-bandorên hêzdar, wek ku Nrf2-hilberên dermanan yên ku îro tê bikaranîn.
Faktoriya nukleer a erythroid 2-related factor, bi gelemperî wekî Nrf2 tê zanîn tê, ku veguherîna gencê, enzymes û antioxidant bersivên hucreyê parastin hucreyek e. Fakslêdana veguherînek cureyê cureyê protein e ku ji bo DNA-ê çêkirina hilberên taybetî û kîmyewî, bi navnîşên glutathione-an jî GST-ê ve girêdayî ye. Activation of Nrf2 hilberîna proteînên çalak aktîf dike ku pêşniyara antioxidant a hêza xwe nîşan bide ku alîkariya alîkariya tedawiya oxidative.
Dr. Alex Jimenez DC, CCST Insight
Enstîtuya Behind Nrf2 Activation
Gava ku Nrf2-çalakkirina xwarinê ya 2004 ya destpêka destpêkê hate çêkirin, agahdariya herî kêm di derbarê fonksiyonê ya Nrf2 de tê zanîn. Di rojnameyên 200 de li ser wêjeya Nrf2, wekî faktoriya nukleer-wekî 2 an jî NFE2L2, tê zanîn ku lêkolîner û lêkolîner tenê bi dest pê kir ku bersiva antioxidant ya Nrf2 di mammalan de bigirin. Li gorî 2017, li ser vê "master regulator," li ser lêkolînên lêkolînê akademiya 9,300 ve hat çap kirin.
Di rastiyê de, Nrf2 gelek antioxidant enzymes dike ku ji wan re genes, ne ji wan re parastina şertên têkildarî yên ku bi hucrey, organ û dawîn bi organîzasyonan re, di bin mercên tendurustî û rêwîtîk de biparêzin. Li gor vê agahdariya nû ya ku ji lêkolînên lêkolînên akademîk yên çapemenî ve, lêkolîner dikarin niha çêtir bikin Nrf2 diranan.
Ji sala 2007-an de, lêkolînên lêkolînê fonksiyona tevlihev a riya Nrf2 destnîşan kirin. Aktîvatorên Nrf2 hatine dîtin ku faktorên strukturên cihêreng ên di laşê mirovan de teqlîd dikin. Bi van rêyan re, aktîvatorên Nrf2 hatine saz kirin ku şert û mercên guherînê li seranserê hucreyê hîs bikin da ku hevsengiyê bihêlin û bersivê bidin hewcedariyên pêşkeftî yên genan.
Çima Bikaranîna Nrf2-Çalakî?
Ji ber ku aboriyên çalakiya Nrf2-ê bi temenê di organîzîzyon de kêm dibe, guhertin dikarin dest pê bibin. Lêkolînên Lêkolîna Lêkolînan nîşan da ku faktorên Nrf2 di hucreyan de bi temenê kêm dike, nîşan bide zêdebûna zextên oksîdative. Cûreyek cûrbecûr ên zehmî yên mîna egherosclerosis û nexweşiya cardiovascular, gehrîtis, kanser, hûrgelê, cureyê 2 êabetes, hypertension, cataracts, û nexweşiyên Alzheimer û bi nexweşiyên Parkinson dikarin ji ber van guhertinan pêşve bibin. Tedbîrên akademî yên bi van pirsgirêkên tendurustiyê têne dîtin.
Bi kapasîteya hucreyê hilweşîna ku hilberîna Nrf2 çalakvanan zêde dike, Nrf2 diranan dikare alîkarîkirina laşê xwe ya mirovê ku bi bandora bandorên oksîdatîf re li dijî veguherandina alîkariya nû bike. Asayîşên fêrbûnê yên fatûnî, an PUFAs, yek ji yekîneyên oksîdî bi hêsantir in û ew bi taybetî bi zehfî ji zextên belaş ên serbixwe ne. Thiobarbituric acid, an jî TBARS, hilberîna bi temenê zêde dibe, nîşan dide ku rêbazek oksîdative bi hev re di rêbazên Nrf2-rêyên rêbaz de bilind kirin.
Ji hêla biyolojîkî ve, vegirtina genê mekanîzmayek bi rastî hêdî ye, bi gelemperî bi demjimêran hewce dike ku bi rêgezê veguhezîne. Wekî encamek, "gelek enzîm xwedan guheztinên xwe yên ser/veqetandî ne ku dikarin di çend hûrdeman de ji hêla enzîmên cûda yên rêkûpêk ve werin rêve kirin. Lekolînwan pêkhateyên xwedan aktîvatorên Nrf2 pêşve xistine ku vê bingeha zanînê ya aktîfkirinê bikar tînin. Çalakkirina Nrf2 ne tenê ji faktora transkripsiyonê ya Nrf2 pêk tê ku ji astengkerê xwe tê derxistin û berbi navika hucreyê ve diçe, lê di heman demê de bi rêzikên DNA yên taybetî ve girêdayî ye da ku îfadeya genê cytoprotective teşwîq bike, leza ku Nrf2 ji navokê derdixe rê dide.
Fêmkirina prosedûra jêbirinê û aktîvkirina Nrf2 di laşê mirov de rê daye lêkolîneran ku hevberdanên çalakvanên cihêreng ên Nrf2 ava bikin da ku bi modulasyona wê ve refleksa genan pêk bînin. Kombûna bingeha zanînê, digel cûrbecûr lêkolînên lêkolînê yên din, alîkariya hilberîna çalakkerên Nrf2 kir ku wekî pêvekên parêzê bikar bînin. Qada agahdariya me bi pirsgirêkên tenduristiya chiropractîk û spinal ve sînorkirî ye. Ji bo nîqaşkirina mijarê, ji kerema xwe ji Dr. Jimenez bipirsin an bi me re têkilî daynin915-850-0900.
Dr. Alex Jimenez
Pirsgirêk Pirsgirêk: Pevçûnê Gelek Kuştina bêyî Surgery
Painşa çokan semptomek navdar e ku dikare ji ber cûrbecûr birîndarbûn û / an şert û mercên çokan çêbibe, tê de birînên werziş. Gelek yek ji nav laşên mirovahiyê ye ku ev yek ji hestê çar çar hestî, çar ligaments, celebên cûda, du mêran û kartilan e. Li gorî Akademiya Xweseriya Danîmarkî ya Amerîkayê, sedemên gelemperî yên tehlûkê yên herî tehlûkê di nav deverên patellar, pirtûka pirtûştê an jî gumperê jumper, û nexweşiya Osgood-Schlatter hene. Tevî ku tehdîtê çermê pir zêde dibe ku li ser 60 salî di nav zarok û ciwanan de dibe ku dibe ku gefê xwînê. Pêwîstiya RICE di malê de piştî rêbazên RICE tê dermankirin, lêbelê, birîndarên giran ên giran dikarin hewceyê baldarî dermanî, bi lênêrîna chiropractîk re.
Antioxidantên zanistî bi zanistî wekî wekhevî têne navnîşan kirin ku pêvajoya oksîdasyonê di laşê mirovî de sînor dike, heger heke heke bêyî çepkirî, ew dikare radikên azad binirxînin ku dibe ku reaksiyonên gelek zîndanê çêbikin ku dibe ku zirarê hucreyî. Bi kêfxweşiyê, bedenek mirov dikare dikare sazkirina pergala parastina bi vî rengî çêbikin, lêbelê, dema ku cureyên oxygenên reaksiyonî re, an jî ROS re, ne nikarin neutralîzekirin, nerazîbûna piçûkek ku gava bi oxygenê veguhestin, zehmet dibe girêdayî ye .
Ji bo ku berdewamkirina li ser axaftina flame, hilberîna dawiyê ya hilberînerê ku ne bandorkirina ROS, an jî cureyên oxygen reaksiyonê reaktîf, rexne dike, di heman demê de, bedena mirovî bi zewacî bi zewacî tê gotin. Tiştek girîng e ku antioxidant hene ku dikarin bi pirsgirêka pirsgirêkên tenduristî re şer bikin û ev antioxidant dikare glutathione e. Tevî ku di 1889 de hate dîtin, bandora antioxidantê glutathione ji mijarên ku di lêkolînên lêkolînê yên modern de bûye yek bû.
Mamosteyê Antîkxidantan: Glutathione
Substanca bihêz tripeptidek e ku ji sîsteîn, asîdê glutamîk û glîsîn pêş dikeve. Ji ber ku jêhatîbûna wê ye ku laşê mirov li hemberê afirandina radîkalên azad biparêze, glutathione di dawiyê de dikare bibe alîkar ku pergala parastinê ya tendurist pêş bikeve. Çi qewimî Raportên zanistî li 2015, ev biryar hate diyarkirin ku aborî ya glutathione ku bi synoxistî bi peroxedîn û katalan re hucreyên parastina li dijî hîdrojen peroxide dike. Ev formula synergistic li dijî cureyên oxygenê re an jî ROS kar dike. Glutathione, peroxidredin û catalase di zêdebûna hûrgelên malostasîsên girîng ên ku hewce ye ku pêvajoyek girîng a hucreyan, tissues û organên tendurust e.
Additionally, glutathione tevahiya pergala pergalê ya parastinê û fonksiyonê bandora lîmphocyte ya girîng e. Li gorî Daîreya Immunochemistry, bi rasterastkirina asta glutathione di laşê mirovan de dikare reaksiyonên ewlehiyê zêde bibin. Bi awayekî nimûne, du ceribandinên cîhanê yên nebaşkirî diyar kir ku tedawiya dermankirinê ya parastinê-nexweşî-bi N-acetyl-cysteine, an jî NAC, encam dibe, di heman demê de di her du rewşan de, di pêvajoyek girîng de di pêvajoyek piranîya piranî de di pêvajoya piraniya immunolojîk de pêkhatî pêkhatî çalakiya hucreyek sirûştî. N-acetyl-cysteine, an jî NAC, sulfur ji glutathione tê bikaranîn û ew bi materyalên zerzîkî re tê bikaranîn, ku paşê ku di nav laşê mirovan de bête avêtin.
Glutathione jî heye ku ji bo lipoic acid veguherîn û herweha ji bo vitamin C û E veguhestin, ku hewce ne hewce ye ku pêvajoya hin sîstemên pêvajoyê bi rêya radestkirina belaşên belaş azad binivîsin. Li ser lêkolîna lêkolînê li ser PLOS ONE, glutathione bi nexweşiyên bi diyabetes metilus, an jî T2DM, û tuberculosyona mycobacterium. Bi gelemperî, kesên ku bi pergalên parastinê yên qelsî ne pişkek heye ku ji bo tbbarkirina xb, an jî tedawîgariya mycobacterium, nexweşî an infeksiyonê nîşan dide. Ji bilî, Kesên bi Tîpa 2-ê diabetes, an jî T2DM-ê, du du sê caran bêtir ji TB-NXXDM-ê bêtir mirovên TB-ê ne. Lêkolînek lêkolînê jî pêşniyaz kir ku asta asta glutathione di nav macrophages de bêtir ji nexweşiyên ku T2DM re dibe sedema xurtkirina kontrola M.Tb an an infeksiyonê. Ev encam nîşan dide ku di asta glutathione de li nexweşan bi T2DM re dibe sedema derfetên mezin a M. Tb nexweş an an infeksiyonê. Her wiha, girêdayî girêdayî Dietro Ghezzi di Brighton û Dibistana Nexweş a Sussexê de, tedbîrên oksîter dikare dibe sedema xemgîniya pergala pergalê û fonksiyonê nebe.
Bi kêfxweşî, glutathione di hêza xurt û kontrolkirina kontrola girîng dike. Bi awayekî nimûne, glutathione ji bo pergala parastinê di nav sîstema parastinê de, ji bo t-lymphocyte belavbûnê, çalakiya phagocytîk, neutrophilên polymerphonîkolojî, û karkerên dendritîk, ji bo bingehîn e ku ji van antigen-pêşniyarên hûrgelan têne çêkirin. . Ewlekariya hûrgelî ya navendî hene ku di destpêkê de di destpêka endocytîk vesiclesên macrophages û dendritîkên dendritîk de dest pê bikin, ji ber vê yekê, peptîdên piçûk li ser hilberkirina T cellsên antigen-specific yên çalak bikin. Herweha, glutathione bi afirandina çêkirina cytokines re dike, û hewce ye ku ew hilberîna germa germên dendritîk ên biparêzin, ku ji bo parastina pergala intracellular-mîkrokotîkên mîkrobokeriyê girîng e.
N-acetyl-cysteine, an jî NAC, zanistî wekî wekî kevneşopiya glutathione tête, ew antioxîdantek pirrjimar e ku wekî antîxîdantek pîvanek radîkal tê bikaranîn. Ji bo rola xwe ya di rola xwe de hat pejirandin acetaminophen zexta zextê, NAC, an N-acetyl-cysteine, hate diyarkirin ku xwediyê gelek sûdên tenduristî û tenduristiyê ye. Ligor Journal Journal, NAC piştgiriya bersivandina tendurustiya tendurustî ya tendurustî dikare dibe ku dibe ku bandorên meriv û pêşniyarên mirovî bandor bike. Lêkolînên lêkolînê diyar kir ku di jinên ku bi pêşiya pêşdestiya vaginos û bacterial vaginosis, bi rojek 0.6-ê NAC-ê bi hevpeymana pêşniyazê 16 ya ducaniyê li dijî pêşdibistana zindî û çêkirina neonatal çêtir parastin. Di encamê de, bandorên erênî yên NAC-ê li ser avakirina pîşesaziyê jî hat dîtin. Piştî sê deqeyên krîza berdewamî hebû, ji sedî 15 de hilberîna hilberîneran bû, çiqas NAC yekem rola sereke dike ku di avakirina pîşesaziyê de çêtir bike û di dema kedê de tevahî tengahî kêm bikin.
Lekolînwanan her weha kifş kir ku NAC, an jî N-acetil-cysteine, dibe ku sûdê bide wan kesên ku bi sendroma ovarian a polycystic, an PCOS hene. PCOS, sendroma hêkokavî ya polîcystîk, nexweşiyek têkildar bi glandên endokrîn e ku bi qasî 5 û 10 ji sedî ya jinên di temenê hilberînê de bandor dike. Di nexweşên weha de, metirsiyek mezintir heye ku bi sendroma metabolê re rûbirû bimîne, ku li wir karanîna NAC alîkariya vegerandina astên însûlînê û hestiyariya tendurist kir.
Dr. Alex Jimenez's Insight
Glutathione wekî "rêjeya antîxîdîdan" ji ber ku rola sereke ya sereke di destkeftî û parastina tevahiya tenduristiyê û başûra de tête dayîn. Dema ku bedenek mirov dikare destûra xwe ya glutathione, xwarinê xurt, qutbûn, zehf, bêhtir bikaranîna dermanan û / an dermanan, zext, trawmayî, temen, zîndanê û radyasyonê dikare hemû asta xwezayî ya glutathione kêm bikin. Ev dikare di şûna mirovên din ên zirarê ji hucreyê de ji zextên oxaxative, radikên azad, infeksiyonê û kansera bêhtir hesab bikin. Ji ber ku tedawiya Glutathione dikare li ser laşê mirovan mirovan gelek qezenc hene. Bi alternatîfên alternatîf ên alternatîf, wekî wekî lênêrîna chiropractîk, asta glutathione dîsa careke din çêbibe ku çêkirina başbûna xweş bikin.
Di heman demê de, profesyonelên tendurustî pêşnîyar kir ku ji bo bikaranîna glutathione vebijarkên alternatîfên din ên alternatîfên alternatîf ên wekî alternatîf, lênêrîna chiyropractîk, ku ji bo tenduristî û başahiyê baştir bikin. Antioxidants ji bo parastina herî baş û girîng e ku reaksiyonên azad ên radîkal ên ku dibe sedema xetera zirarê an zirarê. Antioxidants hêzek mîna glutathione, wekî berê ji jorê re got, paşê alîkariya pêşxistina van radîkalên azad de û bersivê sîstema parastinê ya tenduristî hebe. Lêkolînên lêkolînê hene ku lênêrîna chiyropractîk dibe ku di vê pêvajoyê de roleke girîng hewl bikin, bi xwezayî di çalakiya antîxîdantan de di nav laşê mirovan de zêde bike. Lênêrîna Chiropractîk bi awayek tedbîrên ewlehî û bandor e ku jihevkirinên spînal û pisîkên pirtûka karanîna karanîna misalignasyon, an jî subluxasyonên rastîn bikar bînin, ji bo ku destûra mirovê xwe bi xwezayî bi karanîna dermanên derman / derman û / an jî astengên ziralî xwe xweş bike.
Di dawiyê de, antîoksîdan taybetmendiyên xweyên biyolojîkî bi saya gelek sûdên tenduristiyê nîşan didin, ku dibe ku nuha ji her demê zêdetir hewce be bi êrişa her ku diçe zêde dibe ya stres, nexweşî û qirêjiya cîhana meya nûjen, ku hemî dibin sedema zirarê û / an zirarê hucreyê . Glutathione û pêşengê wê, NAC, an N-acetyl-cysteine, di warê antîoksîdan de rewşa xweya bihêz nîşan didin. Li gel vebijarkên dermankirinê yên alternatîf, wekî lênihêrîna chiropractic, mirov dikare sûdê ji hemî feydeyên ku ev antioxidantê bihêz pêşkêşî dike bike. Berfirehiya agahdariya me bi chiropractic û hem jî bi birîndarî û mercên stûyê ve sînordar e. Ji bo nîqaşkirina mijarê, ji kerema xwe ji Dr. Jimenez bipirsin an jî bi me re têkilî bikin at 915-850-0900.
Dr. Alex Jimenez
Pirsên din
Êşa piştê yek ji sedemên herî zêde yên seqet û rojên ku di xebata cîhanê de cîhanê winda bûne ye. Ji ber ku rastiyek, êşê paşê wekî duyemîn herî gelemperî ji bo serdana doktorê doktora doktorê, tenê ji hêla infeksiyonên bilind-respirasyonê ve tête kirin. Ji sedî 80 ê nifûsa wê di tevahiya jiyana xwe de bi kêmîrek paşa paşê tecrubeyê dike. Spine avahiyek paqij e ku ji hestî, hevpeyman, ligaments û masûlkan, di nav nermên din ên nerm. Ji ber vê yekê, şert û / an jî mercên aggravated, wekî dîsên herniated, dibe ku dibe ku nîşanên nîşan bide paşê paşê. Girtîgehên werzişî yan birîndarên otobloyê gelek caran ji sedemên paşê paşê paşê têne têne, lê carinan, carinan hêsantirên tevgeran dikarin encamên xweş bibin. Bi dilsoz, alternatîfên alternatîf ên alternatîf, wekî lênêrîna chiropractîk, dikare dikare bi alîkariya karanîna spînal û pisîkên pisîkê, dibe ku bi xilasiya xirabtir dibe baştir bike.
Zanîngeha Chiropractor Dr. Alexander Jimenez li çavê xwe dibîne stress, ev çi ye, ew çawa çawa bandor dike û parastina antîxidîdant çawa ji bo çareserkirina rewşê.
Esra Birben PhD, 1 Umit Murat Sahiner MD, 1 Cansin Sackesen MD, 1 Serpil Erzurum MD, 2 û Omer Kalayci, MD1
Kurt: Cureyên oksîjenê yên reaktîf (ROS) ji hêla organîzmayên zindî ve wekî encama metabolîzma xaneyê ya normal û faktorên hawîrdorê, wek qirêjên hewayê an dûmana cixareyê, têne hilberandin. ROS molekulên pir reaktîf in û dikarin strukturên şaneyê yên wekî karbohîdartan, asîdên nukleîk, lîpîd û proteînan zirarê bikin û fonksiyonên wan biguhezînin. Guhertina hevsengiya di navbera oksîdan û antîoksîdan de di berjewendiya oksîdantan de wekî "stresa oksîdasyonê" tê binavkirin. Rêzkirina rewşa kêmkirin û oksîdan (redox) ji bo zindîbûna şaneyê, aktîvkirin, belavbûn û fonksiyona organan krîtîk e. Organîzmayên aerobîk pergalên antîoksîdan ên yekbûyî hene, ku di nav wan de antîoksîdanên enzîmatîk û ne-enzîmatîk hene ku bi gelemperî di astengkirina bandorên zirardar ên ROS de bi bandor in. Lêbelê, di şert û mercên patholojîkî de, pergalên antîoksîdan dikare bête qewirandin. Stresa oksîdatîf beşdarî gelek şert û nexweşiyên patholojîkî dibe, di nav de penceşêr, nexweşiyên neurolojîk, atherosclerosis, hîpertansiyon, iskemiya/perfusion, şekir, sendroma tengasiya nefesê ya akût, fibroza pişikê ya idiopathîk, nexweşiya pişikê ya kronîk a astengdar, û astim. Di vê vekolînê de, em pergalên oksîdan û antîoksîdan ên hucreyî kurt dikin û bandorên hucreyî û mekanîzmayên stresa oksîdative nîqaş dikin.
Key words: antioxidant, oxixant, stressed oxoxative, cureyên oxygen reaksiyonê
(WAO Journal 2012; 5:9-19)
Cureyên oksîjenê yên reaktîf (ROS) ji hêla organîzmayên zindî ve wekî encama metabolîzma xaneyê ya normal têne hilberandin. Di tansiyonên kêm û nerm de, ew di pêvajoyên hucreyên fîzyolojîk de tevdigerin, lê di tansiyonên bilind de, ew guhertinên neyînî li pêkhateyên şaneyê, wek lîpîd, proteîn, û DNA çêdikin. Stresa oksîdatîf tê binavkirin. Stresa oksîdatîf dibe sedema gelek şert û mercên patholojîkî, di nav de kansera, nexweşiyên neurolojîk, 1-6 atherosclerosis, hîpertansiyon, ishemi/perfusion, 7-10 şekir, sendroma tengahiya nefesê ya akût, fibroza pişikê ya îdiopatîk, nexweşiya kronîk a astengdar. ,11 û astimê.14-15 Zîndewerên aerobîk xwedî sîstemên antîoksîdan ên yekbûyî ne, ku di nav wan de antîoksîdanên enzîmatîk û neenzîmatîk hene ku bi gelemperî di astengkirina bandorên zirardar ên ROS de bi bandor in. Lêbelê, di şert û mercên patholojîkî de, pergalên antîoksîdan dikare bête qewirandin. Di vê vekolînê de, em pergalên oksîdan û antîoksîdan ên hucreyî û rêziknameya rewşa kêmkirin û oksîdan (redox) di rewşên tenduristî û nexweşiyê de bi kurtî vedibêjin.
OXIDANTS
Çavkaniyên Navdar ên ROS
ROS ji oxygenê molecular ji ber encamên metabolîzma hucreyek hucreyê hilberandin. ROS dikare di komên 2 de parçe bibin: radîkal û neadadîpên serbixwe. Molekulên ku elektronîk bêpeyalkirî ne û bi vî rengî reklamasyonê bi molekûl re têne radîkal azad kirin. Gava 2 radikên serbixwe elektronên xwe yên bêpeyman ve parve dikin, formên nenadîparêz têne afirandin. Rêzeya 3 ROS e ku girîngiya fîzîkîolojîk anion (o22.), Hydroxyl radîkal (OH), û hîdrojen per peroxide (H2O2). ROS-ê di Table 1 de tête kirin.
Aniona superoksît bi zêdekirina 1 elektron li oksîjena molekulî çê dibe.22 Ev pêvajo bi nîkotîn adenîn dinukleotîd fosfat [NAD(P)H] oxidase an xanthine oxidase an jî bi pergala veguheztina elektron a mîtokondrî ve tê meşandin. Cihê sereke ji bo hilberîna anion superoksîtê mitokondria ye, makîneya şaneyê ku adenosine triphosphate hilberîne. Bi gelemperî, elektron ji bo kêmkirina oksîjenê berbi avê ve bi zincîra veguheztina elektronên mîtokondrî ve têne veguheztin, lê bi qasî 1 heya 3% ji hemî elektronan ji pergalê diherikin û superoksîdê hilberînin. NAD(P)H oxidase di leukocytes, monocytes, û macrofages de polymorphonuclear tê dîtin. Li ser fagosîtozê, van hucreyan teqînek superoksîdê çêdikin ku dibe sedema çalakiya bakterîkujî. Superoksît bi çalakiya superoxide dismutases (SODs, EC 1.15.1.1) vediguhere hîdrojen peroksîtê. Hîdrojen peroksîtê bi hêsanî li ser membrana plazmayê belav dibe. Hîdrojen peroksîtê jî ji hêla xanthine oxidase, amino acid oxidase, û NAD(P)H oxidase-23,24 û di peroxisomes de ji hêla vexwarina oksîjena molekularî ve di reaksiyonên metabolîk de tê hilberandin. Di pey hev reaksiyonên bi navê reaksiyonên Haber Weiss û Fenton de, H2O2 di hebûna metalên veguhêz ên mîna Fe2 an Cu21 de dikare bibe OH21.25.
Fe31 +�.O2�?Fe2 +�O2 Haber Weiss
Fe2 +�H2O2�?Fe3 +�OH�+ .OH Reaksiyona Fenton
O 2 bi xwe jî dikare bi H2 O2 re reaksiyonê bike û OH çêbike. 26,27 Radîkala hîdroksîl ya herî reaktîf a ROS-ê ye û dikare zirarê bide proteîn, lîpîd, û karbohîdartan û DNA. Di heman demê de ew dikare bi girtina elektronek ji asîdên rûn ên polyunsaturated dest bi peroksîdana lîpîdê bike.
Enzîmên granulocîtîk bi rêya eosinophil peroxidase û myeloperoxidase (MPO) reaktîviya H2O2 bêtir berfireh dikin. Di neutrofîlên aktîfkirî de, H2O2 ji hêla MPO ve tê vexwarin. Di hebûna îyona klorîdê de, H2O2 vediguhere asîda hîpoklorê (HOCl). HOCl pir oksîdatîf e û di kuştina pathogenên di rêyên hewayê de rolek girîng dilîze.28 Lêbelê, HOCl dikare bi DNA re reaksiyonê jî bike û danûstendinên DNA-proteînê çêbike û hilberên oksîdasyona pyrimidine hilberîne û klorîdê li bazên DNA zêde bike.29,30 Eosinophil peroxidase. û MPO di heman demê de bi guheztina proteînan bi halojenasyon, nîtrasyon û girêdanên xaçerê yên proteîn ên bi riya radîkalên tîrosîl ve beşdarî stresa oksîdatîf dibe.31
Hêzên azad ên oxgenî yên azad ên peroxyl (ROO $) ne. Forma herî hêsan a radîkayên radîkal-peroxyl (HOO $) ye û roleke di peroxîdasyonê ya fatty ac. Radikên Azad ên dikarin reaksiyonên peroxîdasyonê ji hêla hilberîna methylene karker a hîdrojen atom vebigire. Piştgiriya radîkal paşê bi oxygen re re reaksiyon dike ku radestkirina peroxyl radîkal. Peroxyl radîkal re zencîrekekek destpê dike û pişkên ache yên pîvanên lîberîdsaturated li lîpîdê hîdroperoxides veguherîne. Lipid hîdroşoxoxên pir neheq in û bi hêsanên berhemên duyemîn derxînin, wekî aldehydes (wek 4-hydroxy-2,3-nonenal) û malondialdehydes (MDAs). Isoprostanes ji ber hilberên peroxîdasyonê yên din ên ku ji hêla peroxîdasyonên arachidonic ve têne çêkirin têne çêkirin û di nav plasmma û pêhnavê de kişandine asthmatics .34,35 Peroxidation of lipids integrality of cell membranes distîne û bi rêvekirina membranê membrane dibe .
Hydrogen peroxide, superoxide radîkal, glutathione (GSSG), MDAs, asoprostanes, carbonyls û nitrotyrosine, bi hêsanî ji hêla plomazê, xwîn, an jî bronchoalveolar nimûneyên biyomarker ên bi oxidasyonê bi hêla standardên standardî ve bêne nirxandin.
Çavkaniya Exogenous Çavkanî
Smoke Cigarette
Dûmana cixarê gelek oksîdentan û rakûlên serbixwe û amûrên organîk ên wek oxyoxide û nitrok nitrox .36 Di vê de, bilî kişandina cixarê cixarê cixarê jî di heman demê de mekanîzmayên endogenous jî, wekhevkirina neutrofîl û macrophages, .
Exposure Ozone
Hişyariya Ozone dikare dibe sedema peroxîdê lipid û zêdebûna neutrophilan li seranserê epithelium. Pêwîstina demokrasî ya ozone jî dibe sedema sedemên mediatoran, wekî MPO, protein, kodîkên cationic û herweha dehydrogenase û albînrogenase û albînografîk jî jî albumin .37 Her weha di mijarên tendurustî de, li oozê ozone dihêle sedema karûbarên pulmonaryiyê dike. 38 Cho û al39 diyar kir ku mijara kişandinê (tevlîhevkirina kûrên berbiçav û li germên şewitandinê yên li germê vekirî) catalyoz dide kêmkirina oxygen.
Hyperoxia
Hîperoxia ji bo mercên oxygenê yên ji alozên germî an tissuesên din ên din ên ji asta oksîjenê bilindtir ji asta dermana nermalayî re dikeve. Ew dibe sedema hilberîna oxygenê û cureyên nitrogen
Radiation Ionizing
Radyasyona yonîzasyon, di hebûna O2 de, radîkalên hîdroksîl, superoksît û radîkalên organîk vediguherîne hîdrojen peroksîtê û hîdroperoksîdên organîk. Van celebên hîdroperoksîdê bi îyonên metalê yên çalak ên redox, wek Fe û Cu, reaksiyonên reaksiyonên Fenton re têkildar dikin û bi vî rengî stresa oksîtasyonê derdixin holê.42,43 Narayanan et al44 destnîşan kirin ku fîbroblastên ku ji pariyên alfa re hatine rûxandin di O2 2. û H2O2 ya hundurîn de zêdebûnek girîng heye. hilberîna bi navgîniya NADPH-oksîdaza ku bi parzûna plazmayê ve girêdayî ye.44 Molekulên veguheztina sînyalê, wek kinase 1 û 2-ya ku bi sînyala derveyî hucreyî verastkirî (ERK1/2), c-Jun N-terminal kinase (JNK), û p38, û faktorên veguheztinê, wek aktîvator proteîna-1 (AP-1), faktora nukleer-kB (NF-kB), û p53, têne aktîvkirin, ku di encamê de genên girêdayî bersiva tîrêjê derdixin holê. 45-50 Fotonên Ultraviolet A (UVA) reaksiyonên oksîdative çêdikin. bi heyecana fotosensitîzatorên endogenous, wek porphyrins, NADPH oxidase, û riboflavins. 8-Oxo-7,8-dihydroguanine (8-oxoGua) hilbera sereke ya oksîdasyona DNA-yê bi navbeynkariya UVA-yê ye ku ji oksîdasyona radîkalên OH, oksîjenên 1-elektronî, û oksîjena yekalî pêk tê ku bi giranî bi guanînê re reaksiyonê dike.51 Çêbûna guanînê. Katyonên radîkal ên di DNAya veqetandî de hate destnîşan kirin ku bi bandoriya rasterast a tîrêjên yonîzasyonê pêk tê.52,53 Piştî rûdana tîrêjê yonîzasyon, asta glutathione (GSH) di nav hucreyan de ji bo demek kurt kêm dibe lê dûv re dîsa zêde dibe.54
Heavy Metal Ions
Dikare dikarin zehfên razîk ên reaksiyonên reaksiyonî yên ku ji hêla çalakiya enzyme ve bi lîpîd peroxidasyon û reaksiyonên bi protein û DNA-nNXX-ê re dibe ku hilberên radîkal re hilberînin.
Yek ji mekanîzmayên herî girîng ên hilberîna radîkalên azad ên bi navbeynkariya metal bi reaksiyonek Fenton-ê ye. Iyona superoksît û peroksîta hîdrojenê dikarin bi metalên veguhêz, wek hesin û sifir, bi reaksiyona Haber Weiss/Fenton a katalîzekirî ya metal re têkilî daynin da ku radîkalên OH çêbikin.
Ji xeynî mekanîzmayên celebê Fenton û Haber Weiss-ê, hin îyonên metal dikarin rasterast bi molekulên şaneyê re reaksiyonê bikin da ku radîkalên azad, wekî radîkalên tîolê, hilberînin, an jî rêyên îşaretê yên hucreyê derxînin. Dibe ku ev radîkal bi molekulên tiolê yên din re reaksiyonê bikin ku O22.. O22 çêbikin. vediguhere H2O2, ku dibe sedema hilberîna radîkalên oksîjenê yên zêde. Hin metal, wek arsenît, bi aktîvkirina pergalên hilberîna radîkal ên di hucreyan de nerasterast avakirina ROS-ê çêdikin.56
Arsenîk hêmanek pir jehrî ye ku cûrbecûr ROS-ê çêdike, di nav de superoksît (O2 2), oksîjena yekalî (1O2), radîkal peroksîl (ROO), oksîdê nîtrîk (NO), peroksîdê hîdrojenê (H2O2), û radîkalên peroksîlê dimethylarsinic [( CH3)2AsOO ].57º59 pêkhateyên arsenîk (III) dikarin enzîmên antîoksîdan asteng bikin, nemaze enzîmên girêdayî GSH, wek glutathione-S-transferases (GSTs), glutathione peroxidase (GSH-Px), û GSH reductase, bi girêdanê. - bi komên wan ên sulfhîdrolî (�SH) re.60,61
Lead zêdebûna lipid peroxidation .62 Di nav çalakiya tîska SOD û GP GP-ê de piştî raporkirina rêberê agahdar kirine girîng e .63,64 Replacement of zinc, ku ji bo gelek enzîmên ji bo rêberê komeleya rêfxweş dibe, dibe sedema enzymes. Leşkerkirina rêberê dibe ku ji hêla GST'ê ve bi bandora tissue bandor dike.
ROS-ê ji hêla reaksiyonên kîtal-catalyzed vekirî têne çêkirin-dikare eger binesên DNA-ê. Reterên sê bingehîn, G / C, G / T, û C / T, dibe ku encamên zirarên oxuxative yên bi metal ions, wekî Fe21, Cu21, û Ni21. Reid et al65 diyar kir ku G / C bi gelemperî bi Fe21 hate hilberandin, lê dema veguherîna C / T ji aliyê Cu21 û Ni21 ve hate afirandin.
ANTIOXIDANTS
Laşê mirov bi celebên antioxidant têne çêkirin ku ji bo bandorkirina bandora oxixantan re xizmetê dike. Ji bo armancên tevayî pratîk, ew kategoriyên 2 parçe bibin: enzymatic (Table 2) û neenzymatic (Table 3).
Antioxîdantên Enzymatic
Enzymedîkên antzexîdant ên lûksên SODs (EC 1.15.1.11), catalase (EC 1.11.1.6), û GSH-Px (EC 1.11.1.9) têne. Ji bilî van enzymes, antîxîdîdên din, hebên oxygenase-xNUMX (EC 1), û protein-redox, wekî thioredoxins (TRXs, EC 1.14.99.3), peroxiredoxins (PRXs, EC 1.8.4.10), û glutaredoxins jî hema jî hatine dîtin Parastina antîkxîdantan di pelbên girîng de lîstin.
Ji ber ku superoxide ROS-yê yek ji çavkaniyên hilberîner e, Rûdawiya wê SOD ji bo her hucreyek girîng e. Hemî 3 formên SOD, ku ew, CuZn-SOD, Mn-SOD, û EC-SOD, pir di nav laşê mirovan de tê gotin. Mn-SOD di çarçoveya matrixona mitokondria de ye. EC-SOD di sereke de li matrixa extraellularî, bi taybetî li deverên ku bi mûzîka mîkrojenê de û ezmûnên pişk û pişkên sîstemî yên herêmî tête. Di heman demê de, li ser epithelium, alveolar epithelium û alveolar macrophages .66,67 Bi tevahî, CuZn-SOD û M-SOD têne fikirîn ku fakêşeyên superoxide radîkal wekhev bikin. Di asta herî zêde ya EC-SOD de bi pişkek bi taybetmendiya taybetî ya mîkrofona matrixê ve dibe ku beşek parastina bingehîn ya lîma matrix nîşan bide. 68
H2O2 ku ji aliyê çalakiya SOD ve an çalakiya oxixesên hilberandin, wekî xanthine oxidase, bi avêtina catalase û GSH-Px kêm dibe. Catalase heye ku wekî tetra-mer tête ku ji hêla xenomên wekhev ên 4 ve ye, her yek ji grûpê heme di malpera çalak de digire. Degradation of H2O2 bi guhartina di navbera 2 de peymanên catalase-ferricatalase (avê ve girêdayî avê) û komirê (Istikek bi bi oxygen atom ve girêdayî ye). Catalase jî ji bo kêmkirinahevkirina oksîdatîk ya enzyme (avakirina II), ji hêla H2O2 ve dibe ku ji bo avê kêm dibe .69
Enzymes in di çarçoveyê de kêmkirina H2O2 û lîpîdên hîdroperoxides (ji ber encameke encam a peroxîdasyonê lîpîdê), GSH-Pxs.70 hene GSH-Pxs jî malbata tetrameric enzyme hene ku di hundirê amino acid de selenocysteine de malperên çalak û çalakvanên kêm-molecular-avêtin bikar bînin, wekî GSH, ji bo H2O2 û lîpîd peroxîdên bi alkolên wan re kêm bikin. GSH-PX-1 (cellular GSH-Px) ji hêla þewitandî ye û H2O2 û pîvanên fatîlyayê yên fatixîd kêm dike, lê belê ne esterîtê peroxî lîpîdên ne. -Px-71 (Phospholipid Hîdrobatoxide GSH-Px), ku dikare gelek cûda kêm-molecular-tîrêj bikaribin bikaribin wekheviyên kêm bikin. GSH-Px-4 (GSH-Gastroestestinal GSH-Px) di hucreyên epithelal ên herêmî de tête kirin ku ew kar dike ku xweya xwarinê peroxîdên kêm bikin. 2 GSH-PX-72 (endamê GSH-Px) tenê malbata GSH-Px malbata ku di tête binçavkirin û tête bawerkirin ku yek ji hêla antzxidant anzymeqê di mammalsan de yek ji navdar a pir girîng e. Ji van yekê, GSH-Px-ê pirrjimara pirrfireh di nav lûqa mirovan de lêkolîn e. 3
Herweha, ji bo H2O2 ve ve girêdayî gelek heiol-enzîmên nêzîkî girêdayî, TRXs (TRX1 û TRX2), thioredoxin kêmkirin (EC 1.8.1.9) (TRRs), PRXs (yên thioredoxin peroxidase), û glutaredoxins.74
Du TRXs û TRRs di hucreyên mirovî de, di cîtosol û mîkochondria de hene. Di lenga, TRX û TRR de di bronçalial û alveolar de û macrophages de alveolar têne nîşandan. Di nav hucreyên mirovan de, şeş cûda PRX ve hatine dîtin, di çarçoveya navendî de bêtezirandin. Lêkolînên pisporê girîngiya PRX VI ve di parastina epîtemalaya alveolar de diyar kir. Lûqa Mirov di hemî PRXs de di pergala epithelium, alveolar epithelium û macrophages de .75 PRX V hatiye çêkirin ku di demeke dawî de perçeyek peroxynitrite, 76 ye ku tê wateya ku ew dikare wekî kampa parastina potansiyona ewlehî ya li ROS-mediated lung .77
Ji bo van antioxidants ji bo hewceyê NADPH wek kêmkirina hevpeyman e. NADPH di qada çalak de parastin û ji hêla TRX û GSH kêmtirîn (EC 1.6.4.2), GSSG bi GSH ve, bi hevrêziya GSH-Pxs re hevpeyman dike. NADPH, bi awayekî ve, bi kêmkirina NADP1 bi glucose-6-phosphate dehydrogenase, yekem û rêjeya kêmkirina enzyme ya rêya fosphate-pathê, di dema guhertina glucose-6-fosphate de 6-phosphogluconolactone. Bi hilberîna NADPH, glucose-6-phosphate dehydrogenase kapîtalek girîng a cytosolê ya GSH (GSH / GSSG) tête ye û ji ber vê yekê, dikare enzyme an antioxidant regulatorek tête dîtin. 78,79
GSTs (EC 2.5.1.18), malbatek din a enzîma antîoksîdan, metabolîtên duyemîn, wekî aldehîdên nerazî, epoksîd, û hîdroperoksîdan neçalak dikin. Sê malbatên sereke yên GST-ê hatine şirove kirin: GST-ya sîtosolîk, GST-ya mîtokondrî, 80,81 û GST-ya mîkrozoma-girêdayî ya membranê ya ku di metabolîzma eicosanoid û GSH-ê de xwedî rolek e.82 Heft çînên GST-ya sîtosolîk di mammaliyan de têne nasîn, ku wekî Alpha, Mu, têne destnîşan kirin. Pi, Sigma, Theta, Omega, û Zeta.83~86 Di şert û mercên bê stres de, çîna Mu û Pi GST bi kînaza Ask1 û JNK re têkilî dikin û van kînazan asteng dikin.87~89 Hatiye nîşandan ku GSTP1 ji hev vediqete. JNK di bersiva stresa oksîdasyonê de.89 GSTP1 di heman demê de bi fizîkî bi PRX VI re têkiliyek dike û dibe sedema vegerandina çalakiya enzîma PRX bi riya glutathionylation proteîna oksîdkirî.90
Antioxîdantên Nonenzymatic
Antioxîdanteyên neenzymatîk hene ku amûrên vîtamîn (vitamins C û E), b-carotene, uric acid, û GSH, bi rêvegirtina (lg-glutamyl-l-cysteinyl-L-glycine) ku thiol ( sulfhydryl) grûp.
Vitamin C (Ascorbic Acid)
Water-soluble vitamin C (ascorbic acid) tête navendên antoxxant-a-radîkal an extracellular-aîdous-phase re radigihandin bi radîkayên oxygen-free. Ew vîdyazên azad ên vitamin E veguhestin vîtamîn. Pîvanên plasma wê nîşan dide ku bi temenê kêm bikin. 91,92
Vitamin E (a-Tocopherol)
E li vitamin Lîtin-soluble tête navxweyî ya navxweyî ya hucreyê ya hucreyê û tê parastina sereke li hember zîvanê oxxant-înducedced. Vitamin E ji bo radîkaya radîkal peroxylê ye, ku di dema dema peroxîdasyonê de tête hilberandin. A-Tocopherol forma herî herî çalak a vîtamîn e û hucreya antîxîdant li ser hucreyê mezin e. Vitamin E bi apoptosê hucreyê kansera kanser û rêbazên radîkal ên serbixwe dike. 93
Glutathione
GSH di her dezgehên hucreyek pir mezin e û antîxîdanteya sereke ye. GSH / GSSG rêjeya biryarneyek mezin a oxidative stress e. GSH di gelek awayên antioxidant nîşan dide .94 Ev bi rêya çalakiya GSH-Px ya hîdrojen û lipid peroxides vexwendin. GSH ji bo H2O û O2 kêm bike ji elektrona H2O2 re dike. GSSG dîsa dîsa di GSH de kêmkirina GSH ya ku NAD (P) H bi kar tîne wekî danserê electron. GSH-Pxs ji bo pêşveçûnê ya hucreyê ji hêla peroxidationê lipid e girîng in. Glutathione kêmtir ji protonsan da ku bi lîberên membranan re bike û ji wan êrîşên oxidant biparêzin. 95
GSH ji bo gelek enoxîmên zehmetî yên wekî GSH-Px û veguhastinê ye ku cofactor e. Ew rola xwe di veguherîna vîtamîn C û E veguherîne formên xwe çalakî. GSH hucreyên apoptosê diparêzin bi rêya pêkanîna proapoptotic û pergala antapoptototic veguherînan bike .94 Ev jî herweha faktorên gelek veguherînan e, wekî AP-1, NF-KB, û Sp-1 çalak dike.
Carotenoids (b-Carotene)
Carotenoids pigments in plants in found. Serî, b-carotene hate dîtin ku bi peroxyl (ROO), hîdroxyl (OH), û superoxide (O22) radîkayan .96 Carotenoids bandorên antioxidant di xweseriya asayî ya kêm oxygen nîşan dide lê dibe ku bandorên oxygenant dikarin bi oxygen xemgîniyê .97 Hemî carotenoids û acosên retinoic (RAs) hem dikarin ji hêla faktorên veguherandina veguherandina veguhastinê ve dike .98 b-Carotene çalakvanên NF-kB çalak dike û tevlîheviya (IL) -6 û taktorê nermroş-gerok-nexşîn. Carotenoids jî apoptosyona hucreyan bandor dikin. Bandorên Antiproliferative ya RA di gelek lêkolînan de nîşan da. Ev bandora RA-ê ji hêla reaksiyonên retinoic acid ve tê navnîş kirin û di nav hucreyên hucreyê de cuda ye. Di hucreyên carcinoma mammaryê de, rekêşeya retinoic acidê nîşan dide ku ji bo girtina sefê, apoptosis, an jî hem jî .99,100
Zordariyeke oksîterê dibe ku tewrê di navbera antioxidants û ROS de ji ber ku kêmkirina xelekên antioxidant an danûstandina ROS. Dema ku zextek oksîtatîf tê çêkirin, hucre hewl dike ku bandorên oksîdant û tevlihevkirina zîndanê ji hêla parastina enzymên encodî, faktorên tirsîn û proteinên stratejîk ên stratejîk. yên biryara girîng ên yên zordarî yên oksîterê di bedenê de. Hilberîna bilind ya ROS-ê di bedenê DNA veguherîne, encamên di guhertina protein û lipîdan de, çalakiya gelek faktorên tundûtûj-aşûvanî, û hilberîna pro-inflammatory û cytokines.
Pirsgirêkên Kişandina Oxxatoriyê li DNA
ROS dikare di çend awayan de guhertin, ku di bin bingehên bingehîn de, yek-an anjî ya dNA-ê de, pîrînîn, pyrimidîn an guhertinên şekir-germ-ên, mutations, hilweşandin an jî wergerandin, û bi pêvajoya pêvajoyê re derbas dibe. Pir ji van guhertoyên DNA (Nig 1) bi pirrjimara carcinogenesis, agir û neurodegenerative, cardiovascular, û nexweşiyên xweya otonomê ne. Dûmanên tutunê, mîkrofeyên rûkoxan, û metrên nonredox, wek iron, cadmium, chrom û arsenîk jî, tevlî carcinogenesis û agir têne çêkirin û ji hêla radîkayên serbixwe an jî bi komên thiol ve girêdayî ye. Formasyona 8-OH-G ji ber zehmetiya oksîterê ya herî baş-naskirî ye, û ji bo carcinogenesis ji bo biomarker potansiyona e.
Herêmên pêşverû yên genan rêzikên lihevkirinê yên ji bo faktorên veguheztinê dihewîne. Van cîhên girêdana faktora transkrîpsiyonê rêzikên GC-dewlemend hene ku ji bo êrîşên oksîdant hesas in. Çêbûna DNA ya 8-OH-G di cihên girêdana faktora transkripsiyonê de dikare girêdana faktorên transkripsiyonê biguhezîne û bi vî rengî vegotina genên têkildar biguhezîne wekî ku ji bo rêzikên armanca AP-1 û Sp-1 hatî destnîşan kirin.103 Ji bilî 8-OH-G, 8,59 -cyclo-29 -deoksyadenosine (cyclo-dA) jî hate destnîşan kirin ku ger di nav qutiyek TATA de cih bigire transkrîpsiyona ji genek raporter a di pergala şaneyê de asteng dike.104 Proteîna girêdayê TATA bi guheztina guheztina ADN-ê dest bi veguheztinê dike. . Girêdana proteîna TATA-girêdanê dibe ku ji hêla hebûna cyclo-dA ve were asteng kirin.
Zordariyeke oksîterê sedema seqetkirina microsatêl (herêmên tije yên kurtîn) herêmên din. Rengoxên metal ên çalak yên Redox-ê, radîkalên hîdroxyl zêdebûna microsatêl zêde bikin. 105 Tevî ku tevlîhevkirina DNA-ê bi hêsantir bi hêsantir bi hêsanî têne vebirin, ji hêla hucreyên diya-dionên dionên dionî yên dNA-ê ve dibe sedema tehlûkê ya girîng be girîng e. 106
Methylation li CpG giravên DNA li DNA girîng e ku mîkrofîzmê ya girîng e ku dibe ku bi şikandina gene. Oxuxation of 5-MeCyt bi xweya 5-XMUMX-OHMeUra) bi rêya dagirkirinê / reaksiyonên oxixation dikare dibe navendên thymine an jî 5-hydroxymethyl cytosine. 5 Di bilêvkirina zelalkirina gene ya modulasyonê, DNA-methylasyon jî jî xuya dike ku rêxistina chromatin bandor dike. 107 Pevçûnên DNA-ê-methylasyonên ku ji hêla êrîşên oxidatîf ên li ser çalakiya DNA re bandor dike.
Pirsgirêkên Xweseriya Xweseriya Li Lipids
ROS dikare peroksîda lîpîdê çêbike û lihevhatina dulaytên lîpîdê yên parzûnê xera bike ku dibe ku receptor û enzîmên girêdayî parzûnê bêçalak bike û permebûna tevnê zêde bike.109 Berhemên peroksîda lîpîdê, wek MDA û aldehîdên ne têrbûyî, dikarin bi çêkirina proteînên xaçê gelek proteînên hucreyî neçalak bikin. -girêdan.110~112 4-Hîdroksî-2-nenenal dibe sedema kêmbûna GSH-ya navxaneyî û hilberîna peroksîdê çêdike, 113,114 receptora faktora mezinbûna epidermal aktîf dike, 115 û hilberîna fîbronectin çêdike. , wekî biyomarkerên nerasterast ên stresa oksîdatîf hatine bikar anîn, û astên zêde di kondensata nefesê de an şilava lavajê ya bronchoalveolar an pişika nexweşên nexweşiya pişikê ya astengdar an jî cixarekêşan de zêde bûne.116-117
Pirsgirêkên Stenbolê yên Oxidative li Proteins
ROS dikare bibe sedema perçebûna zincîra peptîdê, guheztina bareya elektrîkî ya proteînan, girêdana xaçerê ya proteînan, û oksîdasyona asîdên amînî yên taybetî û ji ber vê yekê dibe sedema zêdebûna metirsiya proteolîzê bi hilweşandina proteazên taybetî.120 Bermahiyên sîsteîn û methionine di proteînan de 121 Oksîdasyona komên sulfhydryl an bermahiyên methionine yên proteînan dibe sedema guhertinên konformasyonê, vebûna proteîn û hilweşandinê.8,121-123 Enzîmên ku metalên wan li ser cîhên wan ên çalak an nêzikî wan in, bi taybetî ji oksîdasyona katalîzasyona metalê re hesastir in. Guhertina oksîdatîf a enzîman ji bo astengkirina çalakiyên wan hatiye nîşandan.124,125
Di hin rewşan de, oksiyonên taybetî yên protein dikarin bibin. Ji bo nimûne, methionine dikare o-tyrosine126 methionine sulfoxide127 û phenylalanine oxaxized; komên sulfhydryl dikarin oxidized bibin ku têkiliyên dolphide yên dîmêlîdê çêbikin; komên 128 û carbonyl dikarin di bin zincên proteîn de bêne şandin. Rêjeya Gamma, oksîteya metal-catalyzed, HOCL, û ozone dikare dibe sedema komên carbonyl. 129
Pirsgirêkên Stressa Xweser a Li ser Transfera Signal
ROS dikare îfadeya çend genên ku di veguheztina sînyalê de têkildar in derxîne.1,130 Rêjeyek bilind ji bo GSH/GSSG ji bo parastina şaneyê ji zirara oksîdatîf girîng e. Xerabûna vê rêjeyê dibe sedema aktîvkirina faktorên transkripsiyona hesas redox, wek NF-kB, AP-1, faktora navokî ya hucreyên T-ya aktîfkirî û faktora hîpoksî-çêkirî 1, ku di bersiva înflamatuar de beşdar in. Çalakkirina faktorên veguheztinê bi riya ROS-ê bi kaskadên veguheztina sînyala ku agahdariya ji derve berbi hundurê hucreyê vediguhezîne pêk tê. Receptorên Tyrosine kinase, piraniya receptorên faktora mezinbûnê, wekî receptorê faktora mezinbûnê ya epidermal, receptorê faktora mezinbûna endotelyal a vaskuler, û receptorê ji bo faktora mezinbûnê ya ku ji trombêlê hatî hilberandin, proteîn tyrosine fosfatases, û serine/threonine kinase hedefên ROS ne.131-133 Kînazên bi sînyala birêkûpêkkirî yên derveyî hucreyî, JNK, û p38, ku endamên malbata proteîna kînaza aktîfkirî ya mîtojen in û beşdarî gelek pêvajoyên di hucreyê de, di nav de pirbûn, cudabûn, û apoptoz, jî dikarin ji hêla oksîdantan ve werin rêve kirin.
Di bin şert û mercên stresa oksîtasyonê de, bermahiyên cysteine li cîhê girêdana DNA-ya c-Jun, hin binbeşên AP-1, û kînaza kB-ya astengker di bin S-glutathiolation-ya vegerê de ne. Hat ragihandin ku Glutaredoxin û TRX di rêzikkirina rêyên îşaretkirinê yên hestiyar ên redox de, yên wekî NF-kB û AP-1, proteîna kinase ya p38-mîtojen-aktîfkirî, û JNK de rolek girîng dileyzin.134-137
NF-kB dikare di berteka şert û mercên stresa oksîdatîf de, wek ROS, radîkalên azad, û tîrêjên UV-ê were aktîfkirin.138 Fosforîlasyona IkB NF-kB azad dike û dihêle ku bikeve nav nucleusê da ku veguheztina genê çalak bike.139 Hejmarek kînas hene. hate ragihandin ku IkBs li bermahiyên serine fosforîlate dike. Ev kînaz ji bo aktîvkirina NF-kB hedefên sînyalên oksîdatîf in.140 Ajanên kêmker girêdana ADN-ê ya NF-kB zêde dikin, lê ajanên oksîter girêdana DNA ya NF-kB asteng dikin. TRX dibe ku di rêziknameya NF-kB de 2 kiryarên dijberî pêk bîne: di sîtoplazmayê de, ew hilweşandina IkB asteng dike û aktîvkirina NF-kB asteng dike lê girêdana DNA ya NF-kB di navokê de zêde dike.141 Çalakkirina NF-kB bi hilweşandina girêdayî oksîdasyonê. IkB di çalakkirina gelek genên parastina antîoksîdan de encam dide. NF-kB îfadeya çend genên ku beşdarî bersivdayîna parastinê dibin, wekî IL-1b, IL-6, faktora nekroza tumor-a, IL-8, û çend molekulên adhesionê bi rê ve dibe. cudakirina hucreyan.
AP-1 bi hêla dewleta redox ve tê birêve kirin. Heya ku H2O2, hin iyonên metal dikare dikarin çalakiya AP-1 hilberînin. Gelek bi rêjeya GSH / GSSG bi biryara AP-1 zêdetir dike, dema GSSG li dijî DNA-AP-1.144 DNA ya heterodimer bi kêmkirina cysteine di nav de DNA-binding each of Pro protein, 145, dema ku DNA bi bendava AP-1 dikare di gelek hucreyan de, ji hêla gelek hûrgelan ve têne çêkirin, pêşniyar dike ku ji hêla cysteine veguherîna veguherîna înstîtûfa AP-1 DNA binding.
BİXWÎNE
Stressê Oxxative dikare dikare ji zêdekirina hilberandina ROSê bi reaksiyonên metabolîk ên ku oksîjen bikar bînin û guhdarîbûna navbera navbera oxidant /antioxidant bi helwestên oksîdantiyê hene. ROS ji hêla çalakiyên metabolîk û faktorên hawirdorê ve têne hilberandin, wekî asikên asmanî an cixarê cixarê têne hilberandin. ROS ji molekêşên xwe yên rexnegir ên pirrjimarte in ji ber avahiyên wan di bin hucreyên biyolojîk de bi reaksiyonê re wekî rehberî, wekî wekî carbohydrate, nîjîkên nucleîk, û proteîn, û karên xwe biguherînin. ROS jî behsa behsa gelek genes bandor dike ku ji hêla faktorên veguherînek rexnegir û hestone acetylasyon / deacetilasyonê veguherîna hûrgotinê ya kromîtîk-rexne. Regulasyon ya dewleta redoxî ji bo fêrbûna sela, çalakkirinê, hilweşandinê, û organê rêxistinê ye.
REFERENCES
1. Valko M, Rhodes CJ, Moncol J, Izakovic M, Mazur M. Radîkalên belaş, metal û antîoksîdan di kansera stresê ya oksîdative de. Chem Biol Têkilî. 2006; 160:1:40.
2. Halliwell B, Gutteridge JMC. Radîkal ên di Biology û Medicine de. 3rd ed. New York: Press University of Oxford; 1999.
3. Marnett LJ. Peroksîdkirina lîpîdê û zirara DNA ji hêla malondialdehyde. Mutat Res. 1999;424:83-95.
4. Siems WG, Grune T, Esterbauer H. 4-Damezrandina Hydroxynonenal di dema ischemia û reperfusion roviya piçûk a rat. �Life Sci. 1995;57:785~789.
5. Stadtman ER. Rola cureyên oksîdan di pîrbûnê de. Curr Med Chem. 2004;11:1105-1112.
6. Wang MY, Dhingra K, Hittelman WN, Liehr JG, deAndrade M, Li DH. Zêdekirinên DNA yên malondialdehyde-a ku ji hêla peroksîdasyona lîpîdê ve hatî çêkirin di nav tevnên sîngê mirovan de. Penceşêrê Epidemiol Biomarkers Prev. 1996;5:705~710.
7. Jenner P. Stresa oksîdatîf di nexweşiya Parkinson de. Ann Neurol. 2003; 53: S26-S36.
8. Lyras L, Cairns NJ, Jenner A, Jenner P, Halliwell B. Nirxandina zirara oksîterî ya proteîn, lîpîd û DNA di mêjî de ji nexweşên bi nexweşiya Alzheimer. J Neurochem. 1997;68:2061-2069.
9. Sayre LM, Smith MA, Perry G. Kîmya û biochemistry of stress oxidative di nexweşiya neurodegenerative. Curr Med Chem. 2001;8:721~738.
10. Toshniwal PK, Zarling EJ. Delîl ji bo zêdekirina peroksîdana lîpîdê di skleroza pirjimar de. Neurochem Res. 1992; 17:205-207.
11. Dhalla NS, Temsah RM, Netticadan T. Rola stresa oksîdative di nexweşiyên dil de. J Hypertens. 2000;18:655~673.
12. Kasparova S, Brezova V, Valko M, Horecky J, Mlynarik V, et al. Lêkolîna stresa oksîdative di modelek mişkî ya hîpoperfusyona mêjî ya kronîk de. Neurochem Int. 2005;46:601~611.
13. Kerr S, Brosnan MJ, McIntyre M, Reid JL, Dominiczak AF, Hamilton CA. Hilberîna aniona superoksîdê di modelek hîpertansiyona genetîkî de zêde dibe: rola endothelium. Hîpertansiyon. 1999;33:1353-1358.
14. Kukreja RC, Hesen ML. Pergala radîkal a azad a oksîjenê: ji hevkêşan bi navgîniya danûstendinên membran-proteîn heya birîn û parastinê ya dil. Cardiovasc Res. 1992;26:641~655.
15. Asami S, Manabe H, Miyake J, Tsurudome Y, Hirano T, et al. Kişandina cixareyê di cîhek navendî ya pişika mirovî de dibe sedema zêdebûna zirara DNA ya oksîdative, 8-hydroxydeoxyguanosine. Carcinogenesis. 1997;18:1763-1766.
16. Andreadis AA, Hazen SL, Comhair SA, Erzirom SC. Di astimê de bûyerên oksîdatîf û nîtrosaz. Azad Radic Biol Med. 2003;35:213-225.
17. Comhair SA, Ricci KS, Arroliga M, Lara AR, Dweik RA, et al. Têkiliya kêmbûna pergalî ya superoxide dismutase bi astengkirina hewayê di astimê de. Am J Respir Crit Care Med. 2005; 172:306-313.
18. Comhair SA, Xu W, Ghosh S, Thunnissen FB, Almasan A, et al. Di patofîzyolojiya nûvekirina rêça hewayê astim û reaktîf de neçalakkirina Superoxide dismutase. J Pathol im. 2005;166:663.
19. Dut R, Dizdar EA, Birben E, Sackesen C, Soyer OU, Besler T, Kalayci O. Stresa oksîdatîf û diyarkerên wê di rêyên hewayê yên zarokên bi astimê de. Alerjî. 2008;63:1605-1609.
20. Ercan H, Birben E, Dizdar EA, Keskin O, Karaaslan C, û hwd. Stresa oksîdatîf û diyarkerên genetîkî û epîdemîolojîk ên birîna oksîdantê di astima zaroktiyê de. J Allergy Clin Immunol. 2006;118:1097~1104.
21. Fitzpatrick AM, Teague WG, Holguin F, Yeh M, Brown LA. Bernameya Lêkolînê ya Astimê ya Zindî. Homeostasisa glutathione ya hewayê di zarokên bi astima giran de tê guheztin: delîl ji bo stresa oksîdan. J Allergy Clin Immunol. 2009;123:146~152.
22. Miller DM, Buettner GR, Aust SD. Metalên veguhêz wekî katalîzatorên reaksiyonên "otoksîdasyonê". Azad Radic Biol Med. 1990; 8:95-108.
23. Dupuy C, Virion A, Ohayon R, Kaniewski J, D�me D, Pommier J. Mekanîzmaya avakirina hîdrojen peroksîtê ku ji hêla NADPH oxidase ve di membrana plazmaya tîrîdê de hatî katalîz kirin. J Biol Chem. 1991;266:3739~3743.
24. Granger DN. Rola xanthine oxidase û granulocytes di birîna ischemiareperfusion de. J Physiol im. 1988;255:H1269�H1275.
25. Fenton HJH. Di hebûna hesin de oksîda asîda tartarîk. J Chem Soc. 1984;65:899�910.
26. Haber F, Weiss JJ. Parvekirina katalîtîk a hîdrojen peroksîtê bi xwêyên hesin. Proc R Soc Lond Ser A. 1934;147:332�351.
27. Liochev SI, Fridovich I. Haber Weiss 70 sal şûnda çerx kir: nêrînek alternatîf. Redox Rep. 2002; 7:55~57.
28. Klebanoff SJ. Myeloperoxidase: dost û dijmin. J Leukoc Biol. 2005;77:598.
29. Whiteman M, Jenner A, Halliwell B. Guherandinên bazê yên hîpochlorous asîdê di ADN-ya tîmusê ya golikê veqetandî de. Chem Res Toxicol. 1997;10:1240~1246.
30. Kulcharyk PA, Heinecke JW. Asîda hîpochlorous ku ji hêla pergala myeloperoxidase ya fagocytesên mirovan ve hatî hilberandin, di navbera DNA û proteîn de girêdanên xaça kovalent çêdike. Biyokîmya. 2001;40:3648~3656.
31. Brennan ML, Wu W, Fu X, Shen Z, Song W, et al. Çîrokek ji du nakokiyan: pênasekirina hem rola peroxidases di avakirina nitrotyrosine de di vivo de bi karanîna eosinophil peroxidase û mişkên kêmasiya myeloperoxidase, û hem jî cewhera celebên nîtrojenê reaktîf ên ku ji hêla peroxidase ve têne hilberandin. J Biol Chem. 2002;277:17415~17427.
32. Denzler KL, Borchers MT, Crosby JR, Cieslewicz G, Hines EM, et al. Degranulasyona berfireh a eosinophil û oksîdasyona bi navbeynkariya peroksîdazê ya proteînên rêyên hewayê di modelek ovalbumin-ceribandina mişkî ya iltîhaba pişikê de pêk nayê. J Immunol. 2001;167:1672-1682.
33. van Dalen CJ, Winterbourn CC, Senthilmohan R, Kettle AJ. Nîtrît wekî substrat û astengkerê myeloperoxidase. Encamên ji bo nîtrasyon û hilberîna asîda hîpochlorous li cihên iltîhabayê. J Biol Chem. 2000;275:11638~11644.
34. Wood LG, Fitzgerald DA, Gibson PG, Cooper DM, Garg ML. Peroksîda lîpîdê ya ku ji hêla îsoprostanên plazmayê ve hatî destnîşankirin bi giraniya nexweşiyê di astima sivik de têkildar e. Lipids. 2000; 35: 967 974.
35. Montuschi P, Corradi M, Ciabattoni G, Nightingale J, Kharitonov SA, Barnes PJ. Zêdebûna 8-îsoprostane, nîşanek stresa oksîdative, di kondensata derkirî ya nexweşên astmayê de. Am J Respir Crit Care Med. 1999;160:216-220.
36. Dêra DF, Pryor WA. Kîmya-radîkala azad a dûmana cixareyê û encamên wê yên toksîkolojîk. Perspektîfa Tenduristiya Jîngehê. 1985;64:111~126.
37. Hiltermann JT, Lapperre TS, van Bree L, Steerenberg PA, Brahim JJ, et al. Zehfkirina ozonê ya ku di tîrêjê û şilava şûştina bronkî de ji astiman têne nirxandin: di lêkolînên epîdemîolojîk ên li ser qirêjiya hewayê û astimê de amûrek nû ya nedesthilatdar. Azad Radic Biol Med. 1999;27:1448-1454.
38. Nightingale JA, Rogers DF, Barnes PJ. Bandora ozona hilanîn li ser oksîdê nîtrîkî, fonksiyona pişikê, û pişika vekêşandî di mijarên normal û astim de. Thorax. 1999;54:1061-1069.
39. Cho AK, Sioutas C, Miguel AH, Kumagai Y, Schmitz DA, et al. Çalakiya Redox ya keriyên hewayê li deverên cihêreng ên li Gola Los Angelesê. Environ Res. 2005; 99:40~47.
40. Comhair SA, Thomassen MJ, Erzirom SC. Di rêyên hewayê yên mirovên saxlem de ku ji %2 O(100) an dûmana cixareyê ne, veguheztina cûda ya glutathione peroxidase ya derveyî hucreyî û synthase oxide nitric 2. Am J Respir Cell Mol Biol. 2000;23:350~354.
41. Matthay MA, Geiser T, Matalon S, Ischiroopoulos H. Birîna pişikê ya bi navbeynkariya oksîdantê di sendroma tengahiyê ya tîrêjê de. Crit Care Med. 1999;27:2028-2030.
42. Biaglow JE, Mitchell JB, Held K. Girîngiya peroxide û superoxide di bersiva X-ray de. Int J Radiat Oncol Biol Phys. 1992;22:665~669.
43. Chiu SM, Xue LY, Friedman LR, Oleinick NL. Hesaskirina bi îyona sifir a cîhên girêdana matrixa nukleerî ji tîrêjên ionîzasyonê re. Biyokîmya. 1993;32:6214~6219.
44. Narayanan PK, Goodwin EH, Lehnert BE. Parçeyên alfa di hucreyên mirovan de hilberîna biyolojîkî ya anyonên superoksîdê û hîdrojen peroksîtê didin destpêkirin. Cancer Res. 1997;57:3963~3971.
45. Tuttle SW, Varnes ME, Mitchell JB, Biaglow JE. Hestiyariya ji oksîdantên kîmyewî û radyasyonê di xetên hucreyên CHO de ku di çalakiya çerxa pentozê ya oksîdative de kêm in. Int J Radiat Oncol Biol Phys. 1992; 22: 671.
46. Guo G, Yan-Sanders Y, Lyn-Cook BD, Wang T, Tamae D, et al. Manganese
di radiationinduced de behsa gene expressionê de dermanê superoxide
bersivên adaptive. Mol Cell Biol. 2003;23:2362-2378.
47. Azzam EI, de Toledo SM, Spitz DR, JB Little. Metabolîzm
Modulates veguherandina hûrgelan û avakirina micronucleus di bin rêberê
hucreyên ji kîtîkên fikroblasts yên irradiated. Resor Cancer.
2002; 62: 5436 5442.
48. Leach JK, Van Tuyle G, Lin PS, Schmidt-Ullrich R, Mikkelsen RB.
Ionizing radiation-insulated, generation of mitochondria-dependent reactive
oksîjen/nîtrojen. Cancer Res. 2001;61:3894~3901.
49. Dent P, Yacoub A, Fisher PB, Hagan MP, Grant S. MAPK di rê de
bersivên radyasyonê. Oncogene. 2003;22:5885~5896.
50. Wei SJ, Botero A, Hirota K, Bradbury CM, Markovina S, et al. Thioredoxin
veguheztina navokî û pêwendiya bi faktora redox-1 re di bersiva tîrêjên ionîzasyonê de faktora veguheztina AP-1 çalak dike. Cancer Res. 2000;60:6688~6695.
51. Cadet J, Douki T, Gasparutto D, Ravanat JL. Zirara oksîdatîf a ADNyê: avabûn, pîvandin û taybetmendiyên biyokîmyayî. Mutat Res. 2003;531:5�23.
52. Yokoya A, Cunniffe SM, ONeill P. Bandora hîdrokirinê li ser înduksiona şikestin û birînên bingehê di fîlimên DNAya plasmîdê de bi gamaradîasyonê. J Am Chem Soc. 2002;124:8859~8866.
53. Janssen YM, Van Houten B, Borm PJ, Mossman BT. Bersivên hucre û tevnvîsê ji zirara oksîdasyonê re. Lab Invest. 1993;69:261�274.
54. Iwanaga M, Mori K, Iida T, Urata Y, Matsuo T, et al. Faktora nukleerî kappa B ve girêdayî ye ku ji hêla gama glutamylcysteine synthetase ve bi tîrêjên ionîzasyonê ve di hucreyên glioblastoma mirovî T98G de ye. Azad Radic Biol Med. 1998;24:1256-1268.
55. Stohs SJ, Bagchi D. Mekanîzmayên oksîdatîf di jehrbûna îyonên metal de. Azad Radic Biol Med. 1995; 18:321~336.
56. Leonard SS, Harris GK, Shi X. Metal-induced stress oxidative and transduction signal. Azad Radic Biol Med. 2004;37:1921-1942.
57. Shi H, Shi X, Liu KJ. Mekanîzmaya oksîdatîf a jehra arsenîk û kanserojenê. Mol Cell Biochem. 2004;255:67~78.
58. Pi J, Horiguchi S, Sun Y, Nikaido M, Shimojo N, Hayashi T. Mekanîzmayek potansiyel a ji bo xerakirina damezrandina oksîda nîtrîkî ya ku ji ber dirêjkirina devkî ya dirêj a arsenatê di kevroşkan de çêdibe. Free Radic Biol Med.2003;35:102�113.
59. Rin K, Kawaguchi K, Yamanaka K, Tezuka M, Oku N, Okada S. şkestinên DNAstrand ên ku ji hêla dimethylarsinic acid, metabolîtek arsenîkên neorganîk ve têne çêkirin, ji hêla radîkalên anion superoxide ve bi hêz têne zêdekirin. Biol Pharm Bull. 1995; 18:45.
60. Waalkes MP, Liu J, Ward JM, Diwan LA. Mekanîzmayên di binê kanserojeneya arsenîk de: hestiyariya zêde ya mişkan ku di dema ducaniyê de ji arsenîk a organîk re rû bi rû mane. Toxicology. 2004; 198:31.
61. Schiller CM, Fowler BA, Woods JS. Bandorên arsenîk li ser çalakkirina pyruvate dehydrogenase. Perspektîfa Tenduristiya Jîngehê. 1977; 19:205-207.
62. Monterio HP, Bechara EJH, Abdalla DSP. Tevlêbûna radîkalên azad di porphyriayên neurolojîk û jehrîbûna lîberê de. Mol Cell Biochem. 1991; 103:73~83.
63. Tripathi RM, Raghunath R, Mahapatra S. Xwîn û bandora wê li ser Cd, Cu, Zn, Fe û asta hemoglobînê ya zarokan. Sci Total Environ. 2001;277:161�168.
64. Nehru B, Dua R. Bandora seleniumê parêzê li ser neurotoksîkiya rêber. J Environ Pathol Toxicol Oncol. 1997; 16:47.
65. Reid TM, Feig DI, Loeb LA. Mutagenesis ji hêla radîkalên oksîjenê yên ku ji hêla metal ve têne çêkirin. Perspektîfa Tenduristiya Jîngehê. 1994; 102 (uppl 3): 57~61.
66. Kinnula VL, Crapo JD. Superoxide dismutase di pişikê û nexweşiyên pişika mirov de. Am J Respir Crit Care Med. 2003;167:1600~1619.
67. Kinnula VL. Hilberîn û hilweşandina metabolîtên oksîjenê di dema rewşên înflamatuar de di pişika mirovan de. Curr Drug Alergy Inflamm Armanc dike. 2005;4:465~470.
68. Zelko IN, Mariani TJ, Folz RJ. Malbata multigene ya Superoxide dismutase: berhevokek strukturên genê CuZn-SOD (SOD1), Mn-SOD (SOD2), û EC-SOD (SOD3), pêşkeftin û vegotin. Azad Radic Biol Med. 2002;33:337~349.
69. Kirkman HN, Rolfo M, Ferraris AM, Gaetani GF. Mekanîzmayên parastina katalase ji hêla NADPH ve. Kînetîk û stoichiometry. J Biol Chem. 1999;274:13908~13914.
70. Floh L. Glutathione peroxidase. Jiyana Bingehîn Sci. 1988;49:663~668.
71. Arthur JR. Glutathione peroxidases. Cell Mol Life Sci. 2000;57:1825-1835.
72. Chu FF, Doroshow JH, Esworthy RS. Nîşan, taybetmendî, û belavkirina tevnê ya glutathione peroxidase ya nû ya selenium-girêdayî ya hucreyî, GSHPx-GI. J Biol Chem. 1993;268:2571�2576.
73. Comhair SA, Bhathena PR, Farver C, Thunnissen FB, Erzirom SC. Inductiona glutathione peroxidase ya derveyî hucreyî di pişikên astmatîk de: delîl ji bo rêziknameya redoxê ya vegotinê di hucreyên epîteliya hewaya mirovî de. FASEB J. 2001;15:70~78.
74. Gromer S, Urig S, Becker K. Pergala thioredoxin ji zanistê heya klînîkê. Med Res Rev. 2004;24:40~89.
75. Kinnula VL, Lehtonen S, Kaarteenaho-Wiik R, Lakari E, P�kk� P, et al. Vebêja taybetî ya hucreyê ya peroxiredoxins di pişika mirov û sarkoîdoza pişikê de. Thorax. 2002;57:157~164.
76. Dubuisson M, Vander Stricht D, Clippe A, Etienne F, Nauser T, et al. Peroksîredoksîna mirovî 5 reduktazek peroksînîtrite ye. FEBS Lett. 2004;571:161.
77. Holmgren A. Fonksiyona Antioxidant ya pergalên thioredoxin û glutaredoxin. Signal Redox Antioxide. 2000;2:811~820.
78. Dickinson DA, Forman HJ. Glutathione di parastin û nîşankirinê de: dersên ji tiolek piçûk. Ann NY Acad Sci. 2002;973:488~504.
79. Sies H. Glutathione û rola wê di fonksiyonên hucreyî de. Azad Radic Biol Med. 1999;27:916-921.
80. Ladner JE, Parsons JF, Rife CL, Gilliland GL, Armstrong RN. Rêwiyên pêşveçûnê yên paralel ên ji bo transferazên glutathione: avahî û mekanîzmaya kappa enzîma çîna mîtokondrîal rGSTK1-1. Biyokîmya. 2004;43:52~61.
81. Robinson A, Huttley GA, Booth HS, Lijneya PG. Lêkolînên modelkirin û biyoinformatîk ên glutathione transferaza çîna kappa ya mirovî malbatek sêyemîn a transferase ya nû ya bi homologî bi îzomerazên 2-hîdroksîchromene-2-karboksîlate yên prokaryotî pêşbînî dike. Biochem J. 2004;379:541~552.
82. Jakobsson PJ, Morgenstern R, Mancini J, Ford-Hutchinson A, Persson B. Taybetmendiyên pêkhatî yên hevpar ên MAPEGda supermalbata proteînên têkildar ên membranê yên ku di metabolîzma eicosanoid û glutathione de fonksiyonên pir cihêreng hene. Protein Sci. 1999;8:689�692.
83. Hayes JD, Pulford DJ. Malbata supergene ya glutathione S-transferase: rêziknameya GST û tevkariya isoenzîman di kemoparastina penceşêrê û berxwedana derman de. Crit Rev Biochem Mol Biol. 1995;30:445~600.
84. Armstrong RN. Struktura, mekanîzmaya katalîtîk, û pêşkeftina glutathione transferases. Chem Res Toxicol. 1997; 10:2-18.
85. Hayes JD, McLellan LI. Glutathione û enzîmên girêdayî glutathione parastinek hevrêzkirî ya li dijî stresa oksîdative temsîl dikin. Free Radic Res. 1999;31:273~300.
86. Sheehan D, Meade G, Foley VM, Dowd CA. Struktur, fonksiyon û pêşkeftina glutathione transferases: bandorên ji bo dabeşkirina endamên ne-mammalî yên supermalbatek enzîmek kevnar. Biochem J. 2001;360:1º16.
87. Cho SG, Lee YH, Park HS, Ryoo K, Kang KW, et al. Glutathione S-transferase Mu îşaretên aktîfkirî yên stresê bi tepisandina kînaza rêkûpêk a apoptosis 1 modul dike. J Biol Chem. 2001;276:12749~12755.
88. Dorion S, Lambert H, Landry J. Çalakkirina riya nîşana p38 bi şoka germê veqetandina glutathione S-transferase Mu ji Ask1 vedihewîne. J Biol Chem. 2002;277:30792~30797.
89. Adler V, Yin Z, Fuchs SY, Benezra M, Rosario L, et al. Rêziknameya îşaretkirina JNK ji hêla GSTp. EMBO J. 1999;18:1321~1334.
90. Manevich Y, Feinstein SI, Fisher AB. Çalakkirina enzîma antîoksîdan 1-CYS peroxiredoxin hewce dike ku glutathionylation bi navbeynkariya heterodimerîzasyonê bi pGST ve were kirin. Proc Natl Acad Sci US A. 2004;101:3780~3785.
91. Bunker VW. Radîkalên azad, antîoksîdan û pîrbûn. Med Lab Sci. 1992;49:299�312.
92. Mezzetti A, Lapenna D, Romano F, Costantini F, Pierdomenico SD, et al. Stresa oksîdative ya pergalê û têkiliya wê bi temen û nexweşiyê re. J Am Geriatr Soc. 1996;44:823~827.
93. White E, Shannon JS, Patterson RE. Têkiliya navbera vitamin û
bikaranîna lêzêdekirina kalsiyûmê û kansera kolonê. Penceşêrê Epidemiol Biomarkers Prev. 1997;6:769~774.
94. Masella R, Di Benedetto R, Vari R, Filesi C, Giovannini C. Mekanîzmayên nû yên pêkhateyên antîoksîdan ên xwezayî di pergalên biyolojîkî de: tevlêbûna glutathione û enzîmên girêdayî glutathione. J Nutr Biochem. 2005; 16:577~586.
95. Curello S, Ceconi C, Bigoli C, Ferrari R, Albertini A, Guarnieri C. Guhertinên di rewşa glutathione dil de piştî iskemî û reperfusion. Experientia. 1985;41:42~43.
96. El-Agamey A, Lowe GM, McGarvey DJ, Mortensen A, Phillip DM, Truscott TG. Kîmyaya radîkal a carotenoid û taybetmendiyên antîoksîdan / pro-oxidant. Arch Biochem Biophys. 2004;430:37~48.
97. Rice-Evans CA, Sampson J, Bramley PM, Holloway DE. Çima em li bendê ne ku carotenoîd in vivo bibin antîoksîdan? Free Radic Res. 1997; 26:381.
98. Niles RM. Di kemoparastinê û dermankirina penceşêrê de rêyên nîşankirinê. Mutat Res. 2004;555:81.
99. Donato LJ, Noy N. Tepeserkirina mezinbûna kansera memikan ji hêla retînoîk asîdê ve: genên proapoptotîk ji bo receptorê retînoîk asîdê û îşaretkirina proteîna II-ya şaneya asîda retînoîk a xaneyî hedef in. Cancer Res. 2005;65:8193~8199.
100. Niizuma H, Nakamura Y, Ozaki T, Nakanishi H, Ohira M, et al. Bcl-2 ji bo mirina hucreya apoptotîk a ku di neuroblastoma de ji hêla retinoic acid-ê ve hatî çêkirin vesazkerek bingehîn e. Oncogene. 2006;25:5046~5055.
101. Dalton TP, Shertzer HG, Puga A. Rêzkirina vegotina genê bi oksîjena reaktîf. Ann Rev Pharmacol Toxicol. 1999;39:67-101.
102. Scandalios JG. Bersivên genomîk ên ji stresê oksîdative. In: Meyers RA, weş. Encyclopedia of Molecular Cell Biology and Molecular Medicine. Vol 5. 2nd ed. Weinheim, Almanya: Wiley-VCH; 2004: 489.
103. Ghosh R, Mitchell DL. Bandora zirara DNA-ya oksîdative ya di hêmanên promotor de li ser girêdana faktora transkripsiyonê. Nucleic Acids Res. 1999;27:3213-3218.
104. Marietta C, Gulam H, Brooks PJ. Birînek yek 8, 50-cyclo-20-deoxyadenosine di qutiyek TATA de pêşî li girêdana proteîna girêdana TATA digire û bi tundî veguheztinê di vivo de kêm dike. Tamîrkirina DNA (Amst). 2002;1:967~975.
105. Jackson AL, Chen R, Loeb LA. Induction of instability of microsatellite
bi zirara DNA ya oksîdative. Proc Natl Acad Sci US A. 1998;95:12468~12473.
106. Caldecott KW. Têkiliyên proteîn-proteîn di dema tamîrkirina şkestîna yek-zindî ya ADNya mammalan de. Biochem Soc Trans. 2003;31:247-251.
107. Cooke MS, Evans MD, Dizdaroglu M, Lunec J. Zerara DNA ya Oxidative: mekanîzma, mutasyon, û nexweşî. FASEB J. 2003;17:1195~1214.
108. Jones PL, Wolffe AP. Têkiliyên di navbera rêxistina kromatin û methylasyona DNA de di destnîşankirina vegotina genê de. Semin Cancer Biol. 1999; 9: 339, 347.
109. Girotti AW. Mekanîzmayên peroksîdasyona lîpîdê. J Free Radic Biol Med. 1985; 1:87.
110. Siu GM, Draper HH. Metabolîzma malonaldehyde di vivo û in vitro de. Lipids. 1982; 17:349~355.
111. Esterbauer H, Koller E, Slee RG, Koster JF. Tevlêbûna gengaz a hilbera lîpîd-peroksîdasyonê 4-hîdroksînonenal di avakirina kromolîpîdên fluorescent de. Biochem J. 1986;239:405~409.
112. Hagihara M, Nishigaki I, Maseki M, Yagi K. Guhertinên girêdayî temenê di asta lîpîd peroksîtê de di perçeyên lîpoproteîn ên seruma mirovan de. J Gerontol. 1984;39:269�272.
113. Keller JN, Mark RJ, Bruce AJ, Blanc E, Rothstein JD, et al. 4- Hîdroksînonenal, hilberek aldehîdî ya peroksîda lîpîdê ya parzûnê ye, di sinaptozoman de veguheztina glutamate û fonksiyona mîtokondrîal xirab dike. Neuroscience. 1997;806:85~96.
114. Uchida K, Shiraishi M, Naito Y, Torii Y, Nakamura Y, Osawa T. Çalakkirina rêyên îşaretkirina stresê ji hêla hilbera dawî ya peroksîda lîpîdê ve. 4-hîdroksî-2-nonenal înduktorek potansiyel a hilberîna peroksîdê hundurîn e. J Biol Chem. 1999;274:2234�2242.
115. Suc I, Meilhac O, Lajoie-Mazenc I, Vandaele J, Jurgens G, Salvayre R, Negre-Salvayre A. Çalakkirina receptorê EGF ji hêla LDL-a oksîdekirî ve. FASEB J. 1998;12:665~671.
116. Tsukagoshi H, Kawata T, Shimizu Y, Ishizuka T, Dobashi K, Mori M. 4-Hydroxy-2-nonenal hilberîna fibronectin ji hêla IMR-90 fibroblastên pişikê yên mirovî ve hinekî bi riya aktîvkirina faktora mezinbûna epidermal-girêdayî receptorên derveyî hucreyî zêde dike. rêgeza kinase p44/42 verastkirî. Toxicol Appl Pharmacol. 2002;184:127~135.
117. Montuschi P, Collins JV, Ciabattoni G, Lazzeri N, Corradi M, Kharitonov SA, Barnes PJ. 8-îsoprostane di nexweşên bi COPD û cixarekêşên tendurist de wekî biyomarkerek in vivo ya stresa oksîdative ya pişikê derdixe. Am J Respir Crit Care Med. 2000; 162: 1175-1177.
118. Morrison D, Rahman I, Lannan S, MacNee W. Permebûna epithelial, iltîhaba, û stresa oksîdantê li cîhên hewayê yên cixarekêşan. Am J Respir Crit Care Med. 1999;159:473~479.
119. Nowak D, Kasielski M, Antczak A, Pietras T, Bialasiewicz P. Zêdebûna naveroka madeyên tîobarbiturîk-reaktîf û hîdrojen peroksîtê di kondensata nefesê ya qediya ya nexweşên bi nexweşiya pişikê ya kronîk a domdar a domdar de: bandorek girîng a kişandina cixarê tune. Respir Med. 1999;93:389�396.
120. Kelly FJ, Mudway IS. Oksîdasyona proteînê di navbeyna hewa-pişikê de. Amino Acids. 2003;25:375~396.
121. Dean RT, Roberts CR, Jessup W. Parçekirina polîpeptîdên derveyî û hundurîn hucreyî ji hêla radîkalên belaş ve. Prog Clin Biol Res. 1985;180:341~350.
122. Keck RG. Bikaranîna t-butyl hydroperoxide wekî lêkolînek ji bo oksîdasyona methionine di proteînan de. Anal Biochem. 1996;236:56�62.
123. Davies KJ. Ji hêla radîkalên oksîjenê ve zirar û hilweşîna proteîn. I. Aliyên giştî. J Biol Chem. 1987;262:9895-9901.
124. Stadtman ER. Aks-catalyzed-oksîdasyon-metal-protein-a-metal: mekanîzmolojiya biochemîkî û encamên biyolojîk. Radic Biol Med.
1990; 9: 315 325.
125. Fucci L, Oliver CN, Coon MJ, Stadtman ER. Bêçalakkirina enzîmên metabolîk ên sereke ji hêla reaksiyonên oksîdasyona tevlihev-fonksiyonî ve: bandorek mimkun di zivirandina proteîn û pîrbûnê de. Proc Natl Acad Sci US A. 1983; 80: 1521-1525.
126. Stadtman ER, Moskovitz J, Levine RL. Oksîdkirina bermahiyên methionine yên proteînan: encamên biyolojîkî. Signal Redox Antioxide. 2003;5:577~582.
127. Stadtman ER, Levine RL. Oksîdasyona asîdên amînî yên azad û bermayiyên asîda amînî yên di proteînan de bi navbeynkariya radîkalên belaş. Amino Acids. 2003; 25:207-218.
128. Stadtman ER. Di pîrbûn û nexweşiyên bi temen de oksîdasyona proteînê. Ann NY Acad Sci. 2001;928:22~38.
129. Shacter E. Pîvankirin û girîngiya oksîdasyona proteînê di nimûneyên biyolojîk de. Drug Metab Rev. 2000;32:307�326.
130. Poli G, Leonarduzzi G, Biasi F, Chiarpotto E. Stresa oksîdatîf û nîşankirina hucreyê. Curr Med Chem. 2004;11:1163-1182.
131. Neufeld G, Cohen T, Gengrinovitch S, Poltorak Z. Faktora mezinbûna endothelial vascular (VEGF) û receptorên wê. FASEB J. 1999;13:9�22.
132. Sundaresan M, Yu ZX, Ferrans VJ, Sulciner DJ, Gutkind JS, et al. Rêzkirina hilberîna celebên reaktîf-oksîjenê di fibroblastan de ji hêla Rac1 ve. Biochem J. 1996;318:379~382.
133. Sun T, Oberley LW. Rêziknameya Redox ya aktîvatorên transkrîpsiyonê. Azad Radic Biol Med. 1996;21:335~348.
134. Klatt P, Molina EP, De Lacoba MG, Padilla CA, Martinez-Galesteo E, Barcena JA, Lamas S. Rêzkirina Redox ya c-Jun ADN ve girêdayî bi S-glutathiolation reversible. FASEB J. 1999;13:1481�1490.
135. Reynaert NL, Ckless K, Guala As, Wouters EF, van der Vliet A, Janssen Heininger
YM. Di cîh de vedîtina proteînên S-glutathionylated li dû glutaredoxin-1 katalîzasyona cysteine derivatize. Biochim Biophys Acta. 2006;1760:380~387.
136. Reynaert NL, Wouters EF, Janssen-Heininger YM. Modulasyon ya glutaredoxin-1
îfadeya di modela mişkî ya nexweşiya hewayê alerjîk de. Am J Respir Cell Mol Biol. 2007;36:147~151.
137. Filomeni G, Rotilio G, Ciriolo MR. Nîşana hucreyê û pergala redoksa glutathione. Biochem Pharmacol. 2002;64:1057-1064.
138. Pande V, Ramos MJ. Naskirina molekulê ya 15-deoksîdelta (12,14) prostaglandin J(2) ji hêla faktora nukleer-kappa B û proteînên din ên hucreyî ve. Bioorg Med Chem Lett. 2005;15:4057~4063.
139. Perkins ND. Bi fonksiyona NF-kappaB û IKK re rêyên sînyala hucreyê yek dike. Nat Rev Mol Cell Biol. 2007; 8:49.
140. Gilmore TD. Destpêka NF-kappaB: lîstikvan, rê, perspektîf. Oncogene. 2006;25:6680~6684.
141. Hirota K, Murata M, Sachi Y, Nakamura H, Takeuchi J, Mori K, Yodoi J. Rolên cuda yên thioredoxin di sîtoplazma û di navokê de. Mekanîzmayek du-gavekî ya rêziknameya redoxê ya faktora transkripsiyonê NF-kappaB. J Biol Chem. 1999;274:27891�27897.
142. Ward PA. Rola temamker, kemokîn û sîtokînên birêkûpêk di birîna pişikê ya akût de. Ann NY Acad Sci. 1996;796:104~112.
143. Akira S, Kishimoto A. NF-IL6 û NF-kB di rêziknameya genê cytokine de. Adv Immunol. 1997;65:1:46.
144. Meyer M, Schreck R, Baeuerle PA. H2O2 û antîoksîdan bandorên berevajî li ser aktîvkirina NF-kappa B û AP-1 di hucreyên bêkêmasî de hene: AP-1 wekî faktora duyemîn a antîoksîdan-bersiv. EMBO J. 1993;12:2005-2015.
145. Abate C, Patel L, Rausher FJ, Curran T. Rêzkirina Redox ya çalakiya fos û jun DNA-binding in vitro. Zanist. 1990;249:1157-1161.
146. Galter D, Mihm S, Droge W. Bandorên cihêreng ên glutathione disulphide li ser faktorên veguheztina nukleerî kB û proteîna aktîvator-1. Eur J Biochem. 1994;221:639�648.
147. Hirota K, Matsui M, Iwata S, Nishiyama A, Mori K, Yodoi J. Çalakiya veguherîna AP-1 ji hêla têkiliyek rasterast di navbera thioredoxin û Ref-1 de tê rêve kirin. Proc Natl Acad Sci US A. 1997;94: 3633~3638.
Amûra IFM Find A Practitioner tora referralê ya herî mezin e di Tenduristiya Fonksiyonel de, ku ji bo alîkariya nexweşan pizîşkên Fonksiyonên Fonksiyon li her devera cîhanê ye. IFM Pratîsyenên Destûrkirî di encamên lêgerînê de yekem têne navnîş kirin, ji ber ku perwerdehiya wan a berfireh di Tiba Fonksiyonel de heye
Rezervasyon & Hevdîtinên Serhêl 24/7 *
Pêşkêşvanê Dermanê Karbidest * Hemî Pêşkêşkar di hundurê dadrêsiya qanûnî de dixebitin û qada klînîkî ya pratîkê damezirandiye.*